



Принято считать, что биологическая эффективность ЭМИ определяется его термическим действием, которое достаточно хорошо изучено и широко применяется в технологических процессах и медицине. Однако его взаимодействие со средой не ограничивается тепловым эффектом, так как действие ЭМИ проявляется при малых интенсивностях, когда нагрев тканей не является определяющим или вообще становится невозможным [7]. Актуальность исследований действия слабых ЭМИ определяется резким увеличением количества таких источников ЭМИ, как мобильные и радио-телефоны, настольные компьютеры и ноутбуки, способных уже сейчас оказывать значительное влияние на здоровье человека и окружающий его мир[7], и отсутствием четких и логичных представлений о механизмах их действия на живые системы [9-10]. Ранее было показано, что ЭЭГ и дыхание могут чутко реагировать на действие слабых ЭМИ [20]. Информативность анализа ЭЭГ человека и животных при изучении эффектов ЭМИ и других физических факторов низкой интенсивности была продемонстрирована в ряде недавних публикаций [12,14,16-18]. В литературе имеется немало данных о роли ряда параметров ЭМИ [5-9,13] в изменении функционального состояния животных. Однако вопросы о временных режимах действия ЭМИ и роли таких параметров импульсной модуляции как скважность, длительность и частота следования импульсов, ППЭ импульса до сих пор систематически не исследованы. Но, возможно, именно они в той или иной степени определяют участие резонансных и адаптационных механизмов ЦНС в реакциях на ЭМИ и действие других физических факторов [10,18], а адекватный подбор параметров модуляции и режима воздействий (непрерывного или прерывистого) может быть решающим условием эффективности терапевтических процедур [1].
Наше исследование выполнено с целью изучения динамики изменений электрофизиологических коррелятов функционального состояния кроликов в хронических экспериментах при кратковременном действии слабых ЭМИ: непрерывных без модуляции и с импульсной модуляцией. Изменения спектра ЭЭГ, амплитудно-частотных характеристик дыхания и поведения на включение ЭМИ, действие и в фоне после выключения служили критериями влияния ЭМИ на состояние кро-ликов.
ЭЭГ и другие реакции кроликов регистрировали на полиграфе RM-6000 (Япония) и магнитографе TEAC (Япония) и после оцифровки с частотой дискредитации 102.4 Гц переписывали на диск анализатора сигналов с Фурье-процессором IN-110 (Франция). При первичном анализе ЭЭГ с выделением в ходе опыта отдельных ритмических компонент спектра применяли метод динамического спектрального анализа [2]. Регистрация ЭЭГ, дыхания и ЭКГ в хронических экспериментах на 20 взрослых кроликах-самцах шиншилла осуществлялась посредством электродов и соединительных проводников, изготовленных из графита и карбоновых нитей [4],что позволило разрешить проблему радиопомех [9,15] при действии ЭМИ. Облучение кроликов проводили в безэховой экранированной камере генератором ГСС-12 800 МГц с вертикальным рупорным волноводом. Кролика мягко фиксировали в станке капроновой сеткой так, чтобы длинная ось его тела была перпендикулярна Е-вектору ЭМИ, а голова в 15 см под рупором волновода. Для измерения ППЭ использовали термисторный мост, а его распределение под рупором контролировали с помощью рамочной антенны с термисторным преобразователем. Проведено 2 серии (120 опытов) c непрерывной экспозицией ЭМИ (5 минут в опыте); в 1-й серии (n=30) применяли ЭМИ без модуляции; во 2-й (n=90) ЭМИ с импульсной модуляцией 2, 4, 8, 16 и 32 Гц при скважности 30. Длительность импульсов составляла соответственно 16, 8, 4, 2 и 1 миллисекунды, а ППЭ в импульсе - 15 мВт/см².
Результаты исследования и их обсуждение
На первом этапе обработки полученных записей из суммарных спектров ЭЭГ каждого отведения выделялись спектральные компоненты частотой от 0.2 до 10 Гц, динамика интегральной мощности которых на протяжении опыта изображалась графически для сопоставления с динамикой других реакций кроликов. Пример состояния нормы - рис.1. В норме и фоне до включения ЭМИ у кроликов наблюдаются спонтанные изменения мощности спектра ЭЭГ, особенно характерные для дельта-диапазона (до 4 Гц). При их сопоставлении с динамикой параметров дыхания, частоты сердечных сокращений и поведения было сделано заключение, что такие динамичные изменения спектра ЭЭГ у кроликов отражают регулярную смену периодов бодрствования и дремоты, переходящей в сон. Для определения количественных характеристик спектра ЭЭГ провели для отдельных периодов ЭЭГ гистограммное накопление спектров, полученных при обработке последовательных 5-секундных участков ЭЭГ с перекрытием 50%. Анализ спектров в сопоставлении с другими показателями состояния кроликов позволил выделить у них ряд дискретных состояний, чередующихся в цикле бодрствование-сон. Накопленные спектры ЭЭГ, характерные для каждого состояния (активное и спокойное бодрствование, дремота и поверхностный сон, глубокий сон и пробуждение), представлены на рис. 2. Длительность того или иного состояния варьирует от 10-15 секунд до 3-5 минут и связана с индивидуальными особенностями кроликов. В норме характер состояний индивидуален для каждого животного, что делает проблематичным прямое сравнение статистик опытных серий и нормы, поэтому в дальнейшем изменения каждой реакции на действие ЭМИ в каждом опыте нормировали относительно фоновых показателей до воздействия (в %).
В опытах 1-й серии не обнаружено реакций активации животных на ЭМИ. Анализ изменений спектра ЭЭГ, ЧСС, дыхания в сопоставлением с поведением кроликов показал, что 5-минутная экспозиция ЭМИ не изменяет исходное состояние ни при облучении, ни в фоне после него. В следующих 3-4-х опытах наблюдается постепенное усиление медленной активности в спектре ЭЭГ. Можно заключить, что такие воздействия вызывают в ЦНС животных слабые изменения, наиболее характерные для первого уровня адаптации кроликов [3] или для неспецифической адаптационной реакции «тренировки» [7].
Во 2-й серии уже в первых опытах наблюдали четко выраженные реакции животных на включение и действие ЭМИ. Типичный фрагмент записи реакции кроликов на включение ЭМИ с модуляцией в виде снижения амплитуды ЭЭГ, задержки дыхания (апноэ) с последующим изменением его амплитуды представлен на рис.3. Реакции ЦНС кроликов на модулированное ЭМИ в большой степени зависят от состояния. Включение ЭМИ при активном бодрствовании и глубоком сне не вызывает реакций активации ни в ЭЭГ и дыхании, ни в поведении; а вот в переходных состояниях от бодрствования ко сну и наоборот (таких как дремота и пробуждение) вероятность появления реакции активации достигает 70-80%. Достоверно большее их количество зафиксировано при более
низких частотах модуляции ЭМИ, причем латентный период ON-реакций возрастает с увеличением частоты модуляции.
|
|
|
|
Рис. 1. Динамика изменений спектральной мощности частотных диапазонов ЭЭГ правой зрительной зоны коры головного мозга кролика в норме. I - нижний дельта (до 1 Гц), II - верхний дельта (1-4 Гц), III - тэта (4-7 Гц), IV - альфа (7-10 Гц). По оси X - время опыта в минутах. По оси Y - величина спектральной мощности (отн. ед.) |
Рис. 2. Суммарные спектры ЭЭГ правой зрительной зоны коры головного мозга кроликов в разных функциональных состояниях: А и Б - активное и спокойное бодрствование; В - дремота, Г и Д - разные стадии сна; Е - пробуждение. По оси X - частота спектра в Гц. По Y - величина спектральной мощности (отн. ед.) |
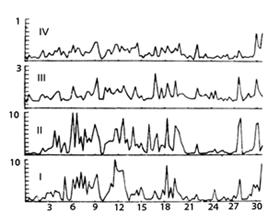
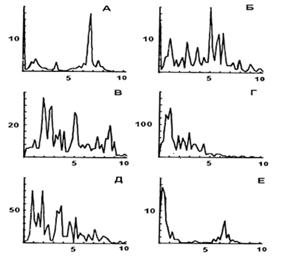
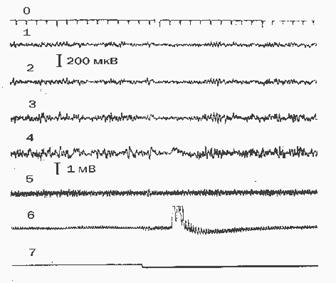
Рис. 3. Фрагмент полиграфической записи электрофизиологических реакций кролика при включении ЭМИ с импульсной модуляцией 8 Гц
0 - отметка времени (секунд); 1- ЭЭГ правой сенсомоторной зоны; 2- ЭЭГ левой сенсомоторной зоны; 3- ЭЭГ правой зрительной зоны; 4- ЭЭГ левой зрительной зоны; 5 - ЭКГ; 6 - пневмограмма; 7 - отметка включения ЭМИ.
Таблица 1. Реакции ЭЭГ в опытах с модуляцией ЭМИ (n - число опытов в серии)
|
|
2-3 Гц (n=20) |
4-5 Гц (n=20) |
8 Гц (n=15) |
16 Гц (n=20) |
32 Гц (n=15) |
|
Количество ON-реакций, % |
65 ± 11 |
55±12 |
33±12 |
25±8 |
13±10 |
|
Латентный период ON-реакций, сек |
1.4±0.2 |
1.8±0.2 |
2.3±0.3 |
2.8±0.4 |
4.0±0.5 |
Среди ON-реакций можно выделить: ориентировочные с латентностью в 1.5-5 секунд и оборонительные - 0.5-1.0 секунд. В поведении кроликов первая проявляется в виде подъема ушей и головы, вторая - в двигательной активности, попытках перекусить провода или убрать голову из-под волновода. При этом в ЭЭГ резко возрастает количество артефактов движения. Такие реакции на действие ЭМИ наблюдались в опытах с модуляцией 2 Гц (15%) и 4 Гц (5%)
Таким образом, ЭМИ с импульсной модуляцией при пятиминутной экспозиции может влиять на функциональное состояние животных. Согласно современным представлениям, основу этого влияния могут составлять реакции тренировки и активации организма в процессах неспецифической адаптации к внешним воздействиям [3, 6, 14] и резонансные механизмы в деятельности ЦНС животных [11-12, 18].
Выводы. В норме у кроликов наблюдается динамичное чередование 6 дискретных состояний в цикле сон/бодрствование, определяемых индивидуальными особенностями животных и имеющих характерные особенности спектра ЭЭГ. Включение ЭМИ без модуляции вызывают в ЦНС кроликов слабые изменения, характерные для неспецифических адаптационных реакций тренировки. Импульсная модуляция ЭМИ резко увеличивает эффективность воздействий и на включение у кроликов регистрируются реакции активации в ЭЭГ и дыхании. Латентные периоды ON-реакций являются почти линейной функцией частоты модуляции и минимальны при модуляции
2-4 Гц.
СПИСОК ЛИТЕРАТУРЫ:
- Боголюбов В.М., Зубкова С.М. // Вопросы курорт. физиотер. леч. физкульт. 1998. № 2 С.3
- Бондарь А.Т., Федотчев А.И. Физиология человека. 1999. Т.25. № 5 С.64
- Васин А.Л., Шафиркин А.В. //Радиобиология. Радиоэкология. 2006. Т. 46 № 4 С.498
- Галеев А.Л., Журавлев Г.И. и др. // Физиол. журн. СССР. 1989. Т. 75 № 12 С.1794
- Гапеев А.Б., Соколов П.А., Чемерис Н.К. // Биофизика 2001. Т. 46 № 4 С.661
- Гаркави Л.Х., Квакина Е.Б., Шихлярова А.И. и др. // Биофизика 1996. Т. 41 №4 С.898
- Григорьев Ю.Г. // Радиобиология. Радиоэкология. 1996. Т. 36 вып.5 С.659
- Григорьев Ю.Г. // Радиобиология. Радиоэкология. 2001. Т. 41 № 5 С.500
- Савин Б.М., Рубцова Н.Б. //Физиология человека и животных Т. 22 Биологическое действие электромагнитных излучений М. 1978. С.68
- Сидоренко В.М. // Биофизика 2001. Т. 46 № 3 С.500
- Федотчев А.И., Бондарь А.Т. //Успехи физиол.наук 1996. Т.27 № 4 С.44
- Федотчев А.И., Бондарь А.Т., Акоев И.Г. // Журн. высш. нервн. деят. 2001. Т. 51 № 1 С.17
- Холодов Ю.А. // Радиобиология. Радиоэкология.1998. Т. 38 № 1 С.121
- Cook C.M., Thomas A.W., Prato F.S. // Bioelectromagnetics. 2002. V. 23 № 2 P.144
- Deboer T. // J. Sleep Res. 1998. V. 7. № 4. P.254
- D´Costa H., Trueman G., Tang L. et al. // Australas Phys. Eng. Sci. Med. 2003. V. 26. № 4 P.162
- Krause C.M., Haarala C., Sillanmaki L. et al. // Bioelectromagnetics 2004. V. 25 № 1 P.33
- Marino A.A., Nilsen E., Frilot C. // Bioelectromagnetics 2003. V. 24 № 5 P.339.
- Salansky N., Fedotchev A., Bondar A. // Neurosci. Biobehav. Rev. 1998. V. 22 № 3 P.395
- Zhuravlev G.I., Fedotchev A.I. J. Biophysics 2005. suppl.2 p.134