



Одной из основных задач науки в современном природопользовании является его переориентация на сохранение и восстановление экологических функций экосистем, в том числе функции поддержания биологического разнообразия. В настоящее время решение этого вопроса может быть эффективно выполнено с позиции популяционной парадигмы организации экосистем [4]. При использовании популяционного подхода экосистема рассматривается как совокупность всех составляющих ее популяций видов разных трофических групп, взаимодействующих между собой на различных уровнях [3, 4, 10]. Оценка сукцессионного статуса лесной экосистемы и потенциала к ее восстановлению в этом случае возможна через оценку состояния популяций древесных растений, как главных эдификаторов в позднесукцессионных сообществах [5]. Целью нашей работы была апробация методики оценки сукцессинного состояния хвойно-широколиственных лесов юга российского Дальнего Востока на основе популяционно-демографического подхода.
Материалы и методы исследования
Для оценки состояния популяции деревьев использован подход, предложенный О.В. Смирновой с соавторами [5], в соответствии с которым вначале закладываются пробные площади определенного размера, где затем проводится перечет древостоя и для каждого дерева указываются его онтогенетическое состояние и уровень жизненности. Полученные данные позволяют построить гистограммы распределения особей по онтогенетическим состояниям (онтогенетические спектры) [5]. На основе анализа характера этих спектров (инвазионные, нормальные, регрессивные) для ранне- и позднесукцессионных древесных растений можно дать оценку сукцессионному статусу сообщества и сделать прогноз его развития. Размер пробных площадей для европейских лесов составляет 0,25–20 га [5]. Для дальневосточных лесов такая работа проведена впервые, а разнообразие видов древесного яруса значительно выше, поэтому нами также проводился подбор оптимального размера пробной площади.
Исследования были проведены на 13 пробных площадях (ПП), расположенных на территории Приморского края (отроги горной системы Сихотэ-Алинь): ПП 2, 3, 4 и 12 находятся на территории Верхнеуссурийского стационара Биолого-почвенного института Дальневосточного отделения РАН (ВУС БПИ ДВО РАН), ПП 1 – в бассейне р. Горная (п. Мартынова поляна), ПП 5 – р. Синегорка (с. Рисовое), ПП 6, 7 и 8 – в бассейне р. Барсуковка, ПП 9, 10 – в бассейне р. Средняя Крыловка и Левая Крыловка (притоки второго порядка р. Уссури), ПП 11 – в бассейне р. Седанка (зеленая зона г. Владивостока), ПП 13 – в бассейне р. Амба (приток первого порядка р. Бикин). Из 13 пробных площадей в типичных кедровниках [1] заложены 7, в южных кедровниках – 4 и в пихтово-еловых лесах – 2.
Предварительный подбор участков для закладки временных пробных площадей осуществлялся на основе данных лесоустройства и анализа космоснимков Landsat ETM + и Spot 5. Для пробной площади выбирался участок древостоя в переделах физиономически выделяемого контура лесной растительности, с относительно однородным рельефом в пределах выбранной экспозиции склона. Исследование пробных площадей проводилось методом сплошного перечета деревьев. При перечете учитывались: вид, возрастное состояние, координаты (с использованием gps-навигаторов). Учет возрастного состояния для растений проводился на основе дискретного описания онтогенеза [8], начиная с виргинильного возрастного состояния. Разделение растений Pinus koraiensis, Picea ajanensis и Abies nephrolepis на возрастные стадии проводилось согласно работам Т.А. Комаровой с соавторами [2, 7], для остальных видов использованы общие принципы описания онтогенеза [9]. Кроме того, на пробной площади отмечались вывально-почвенные комплексы и пни по возможности с указанием вида дерева, его возрастного состояния на момент образования и способа образования пня (естественный облом, выворот дерева или рубка). Возраст рубки определялся ориентировочно по увеличению радиального прироста соседних деревьев или степени разложения послерубочных пней [8]. Давность пожара определялась по наличию долгоживущих деревьев, подпалинам и присутствию крупного валежа [4].
Результаты исследования и их обсуждение
Всего в исследовании использованы материалы 13 пробных площадей, находящихся на разных стадиях восстановительных сукцессий, из них на 9 зафиксированы следы рубок и пожаров (табл. 1), лишь в одном районе (ВУС БПИ ДВО РАН) не обнаружены видимые следы хозяйственной деятельности человека.
Таблица 1
Характеристика пробных площадей
|
№ п/п |
Размер, га |
Общая характеристика |
|
1 |
0,6 |
Кедрово-широколиственный лес с видимыми следами неоднократных рубок (около 10 и 40 лет назад) и пожара (около 200 лет назад) |
|
2 |
1,0 |
Кедрово-широколиственный лес без видимых следов рубки и пожара за последние 350–400 лет |
|
3 |
1,0 |
Елово-пихтовый лес без видимых следов рубки и пожара за 250 лет |
|
4 |
0,6 |
Елово-пихтовый лес вблизи вырубки |
|
5 |
2,0 |
Кедрово-широколиственный лес неоднократно пройден рубками (последние около 20–30 лет назад) |
|
6 |
0,6 |
Подвергавшийся неоднократным рубкам (последней 5–10 лет) и пожарам кедрово-чернопихтово-широколиственный лес |
|
7 |
0,7 |
Кедрово-чернопихтово-широколиственный лес после выборочной рубки (давность последней 10–20 лет) |
|
8 |
0,8 |
Кедрово-чернопихтово-широколиственный лес со следами выборочных рубок (несколько деревьев были срублены 10–20 лет назад) |
|
9 |
0,8 |
Кедрово-широколиственный лес со следами выборочной рубки (давность 30–40 лет) |
|
10 |
0,8 |
Кедрово-широколиственный лес после неоднократной выборочной вырубки, с кратковременным промежутком – последняя рубка около 5 лет, предыдущая ‒ около 25 лет назад |
|
11 |
2,0 |
Кедрово-чернопихтово-широколиственного лес со следами неоднократных рубок и пожаров, давность последней рубки – 30–40 лет |
|
12 |
9,0 |
Кедрово-елово-широколиственный лес без видимых следов рубки и пожаров за последние 300–400 лет |
|
13 |
6,0 |
После пожарный кедрово-широколиственный лес (давность пожара около 250 лет) с наличием следов приисковых рубок, давностью 70–80 лет |
Всего на пробных площадях был встречен 41 вид древесных растений (табл. 2), из них лишь 4 вида отмечены на всех пробных площадях. К ним относятся: Betula costata, Pinus koraiensis, Tilia amurensis, Padus maximowiczii. Два вида (Abies nephrolepis, Acer tegmentosum) присутствовали на 12 пробных площадях из 13. Еще три вида (Acer mono, Fraxinus mandshurica, Ulmus laciniata) отмечены на всех пробных площадях, исключая пробные площади, расположенные в елово-пихтовых лесах (ПП 3 и 4). Наиболее редко встречались Cerasus sargentii, Rhamnus davurica, Betula davurica, B. ermanii, Crataegus pinnatifida, Malus baccata, Padus asiatica, Picea koraiensis. Все перечисленные виды относятся к группе позднесукцессионных. Раннесукцессионные виды, например, Betula platyphylla, всегда малочисленны, если только степень нарушенности древостоя не близка к катастрофической, поэтому в случае смешанных лесов юга российского Дальнего Востока мы не можем использовать для оценки сукцессионного состояния древостоя соотношение числа ранне- и позднесукцессионных видов. Оценка должна основываться на анализе онтогенетических спектров и сравнения их со спектрами видов в ненарушенных сообществах.
Таблица 2
Присутствие видов деревьев на пробных площадях
|
Вид |
Номер пробной площади |
||||||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
|
|
Abies holophylla Maxim. |
+ |
+ |
+ |
+ |
|||||||||
|
Abies nephrolepis (Trautv.) Maxim. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
|
Acer mandshuricum Maxim. |
+ |
+ |
+ |
+ |
+ |
||||||||
|
Acer mono Maxim. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||
|
Acer pseudosieboldianum (Pax.) Kom. |
+ |
+ |
+ |
+ |
+ |
||||||||
|
Acer tegmentosum Maxim. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
|
Acer ukurunduense Trautv. et C.A. Mey. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||||
|
Alnus hirsuta (Spach) Fisch. ex Rupr. |
+ |
+ |
+ |
+ |
|||||||||
|
Betula costata Trautv. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Betula davurica Pall. |
+ |
+ |
|||||||||||
|
Betula ermanii Cham. |
+ |
+ |
|||||||||||
|
Betula platyphylla Sukacz. |
+ |
+ |
+ |
||||||||||
|
Carpinus cordata Blume |
+ |
+ |
+ |
+ |
|||||||||
|
Cerasus sargentii (Rehd.) Pojark. |
+ |
||||||||||||
|
Crataegus pinnatifida Bunge |
+ |
+ |
|||||||||||
|
Fraxinus mandshurica Rupr. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||
|
Fraxinus rhynchophylla Hance |
+ |
+ |
+ |
+ |
|||||||||
|
Juglans mandshurica Maxim. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||||
|
Kalopanax septemlobus (Thunb. ex Murray) Koidz. |
+ |
+ |
+ |
||||||||||
|
Ligustrina amurensis Rupr. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||||
|
Maackia amurensis Rupr. et Maxim. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||||||
|
Malus baccata (L.) Borkh. |
+ |
+ |
|||||||||||
|
Micromeles alnifolia (Siebold et Zucc.) Koehne |
+ |
+ |
+ |
+ |
|||||||||
|
Padus avium Mill. |
+ |
+ |
|||||||||||
|
Padus maackii (Rupr.) Kom. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||||||
|
Padus maximowiczii (Rupr.) Sokolov |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Phellodendron amurense Rupr. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||||
|
Picea ajanensis (Lindl. et Gord.) Fisch. ex Carr. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||||
|
Picea koraiensis Nakai |
+ |
+ |
|||||||||||
|
Pinus koraiensis Siebold et Zucc. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Populus maximowiczii A. Henry |
+ |
+ |
+ |
||||||||||
|
Populus tremula L. |
+ |
+ |
+ |
+ |
|||||||||
|
Quercus mongolica Fisch. ex Ledeb. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|||
|
Rhamnus davurica Pall. |
+ |
||||||||||||
|
Salix caprea L. |
+ |
+ |
+ |
||||||||||
|
Sorbus pochuashanensis (Hance) Hedl. |
+ |
+ |
+ |
+ |
+ |
||||||||
|
Taxus cuspidata Siebold et Zucc. ex Endl. |
+ |
+ |
+ |
||||||||||
|
Tilia amurensis Rupr. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Tilia mandshurica Rupr. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||||
|
Ulmus japonica (Rehd.) Sarg. |
+ |
+ |
+ |
+ |
+ |
+ |
|||||||
|
Ulmus laciniata (Trautv.) Mayr |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||
|
Всего видов: |
25 |
12 |
10 |
10 |
30 |
24 |
25 |
26 |
17 |
16 |
26 |
17 |
23 |
Основываясь на полученных результатах и имеющихся в литературе данных об экологии и биологии видов, мы разделили все учтенные в ходе полевых работ виды на несколько групп (табл. 3).
Таблица 3
Условные группы видов по численности на пробных площадях и размеру территории, необходимой для выявления сукцессионного состояния популяции
|
Группа |
Виды |
|
|
Первый подъярус |
Второй подъярус |
|
|
1. Наиболее многочисленные и долгоживущие виды-эдификаторы смешанных лесов, для которых возможно получить полночленные онтогенетические спектры на пробных площадях размером до 2 га |
Abies holophylla Abies nephrolepis Acer mono Picea ajanensis Pinus koraiensis Tilia amurensis Tilia mandshurica |
Carpinus cordata |
|
2. Виды-эдификаторы, для которых возможно получить полночленные онтогенетические спектры на площади 4–9 га |
Betula costata Fraxinus mandschurica Juglans mandshurica Picea koraiensis Quercus mongolica Ulmus laciniata Ulmus japonica |
Acer pseudosieboldianum Acer tegmentosum Acer ukurunduense |
|
3. Малочисленные (в малонарушенных древостоях) виды, для которых возможно получить полночленные онтогенетические спектры на площади более 9 га |
Acer mandshuricum Alnus hirsuta Betula davurica Betula ermanii Betula mandshurica Fraxinus rhynchophylla Kalopanax septemlobus Maackia amurensis Micromeles alnifolia Phellodendron amurensis Padus avium Populus tremula Populus maximowiczii |
Сerasus maximowiczii Cerasus sargentii Crataegus pinnatifida Ligustrina amurensis Malus mandshurica Padus maackii Salix caprea Sorbus pochuashanensis Rhamnus davurica Taxus cuspidata |
Как показывает анализ изменения онтогенетических спектров с увеличением размера ПП до 2 га, можно получить полночленный онтогенетический спектр для 8 наиболее многочисленных видов-эдификаторов (табл. 3, группа 1). Причем для таких видов, как P. koraiensis, A. nephrolepis, T. amurensis, C. cordata чаще площадь выявления спектра будет составлять от 0,7 га, для остальных видов из этой группы – не менее 2 га. Во вторую группу можно отнести виды, для которых площадь, требуемая для получения полночленного спектра, составляет от четырех до шести гектар. Третья группа видов наиболее многочисленная. Для этих видов характерно небольшое число особей в малонарушенных древостоях и увеличение их численности после нарушения древостоя рубками или пожарами.
В процессе проведения исследований стало ясно, что для большинства видов онтогенетические спектры малоинформативны для анализа сукцессионного состояния древостоя, в связи с тем, что данные по их биологии и экологии отрывочны. Эта работа осложнена тем обстоятельством, что за последние 100–150 лет южная часть российского Дальнего Востока подверглась интенсивному хозяйственному освоению, поэтому для полной оценки сукцессионного состояния лесных сообществ необходимо проводить дальнейшие исследования в малонарушенных древостоях. Тем не менее, основываясь на анализе спектров наиболее многочисленных видов-эдификаторов (первая группа), полученных по материалам временных пробных площадей, а также имеющихся в литературе данных по биологии и экологии видов [2, 6, 7, и др.], мы можем дать характеристику текущей демографической ситуации в древостое и перспективам ее изменений на ближайшие 50–100 лет. Одной из наиболее крупных пробных площадей, заложенной в ненарушенном участке леса, была ПП 12. Полученные на ней материалы были использованы для выявления онтогенетических спектров, характерных для ряда основных видов-эдификаторов (рисунок).
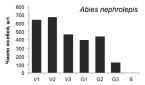
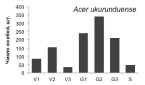
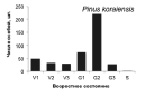
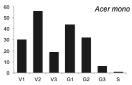
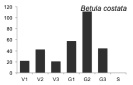
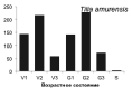
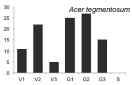
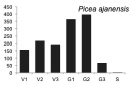

Онтогенетические спектры некоторых древесных видов, характерные для древостоев на поздних стадиях сукцессии. Условные обозначения: V1 – начальная фаза виргинильного возрастного состояния, V2 – промежуточная фаза виргинильного возрастного состояния, V3 – завершающая фаза виргинильного возрастного состояния; G1 – молодое генеративное возрастное состояние, G2 – средневозрастное генеративное возрастное состояние, G3 – старогенеративное возрастное состояние; S – сенильное возрастное состояние
Как видно на рисунке, онтогенетические спектры большинства видов за исключением Abies nephrolepis и Acer mono можно ошибочно отнести к регрессионному типу. Для объяснения этого феномена необходимо учитывать два обстоятельства. Во-первых, в исследовании не учитывались особи в имматурном возрастном состоянии, а во-вторых, на форму спектра имеет значительное влияние продолжительность жизни и скорость прохождения отдельных возрастных состояний каждым видом. Так, например, в спектре P. koraiensis есть большой «провал» в виргинильном возрастном состоянии (v1–v3) (рисунок), но проведенные нами ранее исследования показывают, что у P. koraiensis развитие деревьев во втором пологе древостоя и выход в верхний полог происходит за несколько периодов усиленного роста, чередующихся с замедленным [7]. Таким образом, в одних возрастных состояниях особи длительное время накапливаются, а через другие проходят относительно быстро, поэтому онтогенетический спектр может иметь форму с несколькими вершинами. Подобный вид онтогенетического спектра характерен и для других видов, указанных на рисунке (например, B. costata, A. mono, T. amurensis).
Приведем один пример оценки сукцессионного состояния древостоя по материалам ПП 1. Спектры основных видов-эдификаторов имеют следующий вид: у Pinus koraiensis, Acer mono их можно отнести к инвазионным, у Picea ajanensis и Abies nephrolepis спектры фрагментарные, Tilia amurensis и T. mandshurica также представлены неполными спектрами. Это свидетельствует о том, что древостой находится на раннем этапе сукцессии после среднего нарушения [5] (сложившегося из нескольких слабых – рубок и низового пожара). В ближайшее время значительного изменения видового состава древостоя (при условии отсутствия дальнейших антропогенных нарушений) происходить не будет. В течение этого времени из состава могут исчезнуть лишь раннесукцессионные виды – B. mandshurica, B. davurica, S. caprea, спектры которых на пробной площади фрагментарны и представлены генеративными растениями. Но с учетом того, что у этих видов накапливаются большие запасы жизнеспособных семян в почве, а изреживание древостоя на данном этапе его развития происходит быстрыми темпами, в течение некоторого времени могут образовываться окна в пологе, подходящие для прорастания и развития этих видов.
Заключение
Применение популяционно-демографического подхода в хвойно-широколиственных лесах для оценки сукцессионного состояния возможно, но при этом существует несколько особенностей. Выяснено, что большое число особей раннесукцессионных видов свидетельствует о недавнем нарушении сильной или средней интенсивности, в древостоях, находящихся на поздних стадиях сукцессии, наоборот, число особей этих видов значительно меньше. При этом для анализа онтогенетических спектров важны знания о биологии и экологии видов, поскольку отнесение спектра к тому или иному типу затруднено. Вследствие значительного разнообразия видов древесного яруса для оценки сукцессионного состояния сообщества по онтогенетическим спектрам всех или большей части видов необходима пробная площадь размером не менее 9 га, для оценки только по основным эдификаторам этот размер может быть уменьшен до 2 га.
Рецензенты:
Баркалов В.Ю., д.б.н., заведующий отделом лесных и почвенных ресурсов, Биолого-почвенный институт ДВО РАН, г. Владивосток;
Денисов Н.И., д.б.н., главный научный сотрудник, Ботанический сад-институт ДВО РАН, г. Владивосток.
Работа поступила в редакцию 19.12.2013.
Библиографическая ссылка
Ухваткина О.Н., Омелько А.М. ОЦЕНКА СУКЦЕССИОННОГО СОСТОЯНИЯ ДРЕВОСТОЕВ ХВОЙНО-ШИРОКОЛИСТВЕННЫХ ЛЕСОВ ЮГА РОССИЙСКОГО ДАЛЬНЕГО ВОСТОКА НА ОСНОВЕ ПОПУЛЯЦИОННО-ДЕМОГРАФИЧЕСКОГО ПОДХОДА // Фундаментальные исследования. – 2013. – № 11-5. – С. 948-953;URL: https://fundamental-research.ru/ru/article/view?id=33231 (дата обращения: 20.04.2024).