



Образование активных форм кислорода (АФК), таких как пероксид водорода, супероксидный анион-радикал (О2•–), гидроксильный радикал (ОН–) и синглетный кислород (1О2), происходит в процессе аэробного метаболизма клетки. Окислительный сигнальный каскад запускается в растениях также в ответ на все основные виды стрессоров (чрезмерную или недостаточную освещенность, экстремальную температуру, засуху, засоление, тяжелые металлы, гербициды, ксенобиотики и т.д.). В том случае если антиоксидантная система (АОС) не способна эффективно устранять АФК, в клетке развивается окислительный стресс. Будучи высоко реакционноспособными, АФК могут реагировать со всеми макромолекулами (липидами, нуклеиновыми кислотами, белками, полисахаридами), вызывая их повреждения. Изменения в структуре нуклеиновых кислот, не удаленные репарацией, могут реализоваться в виде генных или хромосомных мутаций. Для защиты от повреждающего действия АФК клетки растений используют различные антиоксидантные системы. К ним относятся неферментативные антиоксиданты, такие как аскорбат, глутатион, токоферол, различные фенольные соединения, а также антиоксидантные ферменты [10, 11].
Цель исследования – определить оксидативный статус кальцефитных растений, произрастающих в различных условиях – на черноземных почвах Воронежской области и в условиях искусственно созданной модели альпийской горки с меловыми почвами.
Материал и методы исследования
В качестве объектов исследования были выбраны такие кальцефитные растения, как Бурачок Гмелина (Alyssum gmelinii), Лен многолетний (Linum perenne), Шлемник приземистый (Scutellaria supina).
Содержание белка в пробах определяли методом Лоури. Соотношение хлорофиллов и каротиноидов (мкг/г сухой массы) вычисляли спектрофотометрически в ацетоновых растительных экстрактах [7].
Пероксидазную активность измеряли по степени окисления о-дианизидина. За единицу ферментативной активности принимали такое количество пероксида водорода (мкмоль), которое разложилось при действии 1 мг пероксидазы за 1 минуту (мкмоль/мин на 1 мг белка) [3].
Определение каталазной активности было основано на реакции разложения пероксида водорода. За единицу активности фермента принимали такое количество (в мкмоль) пероксида водорода, которое разложилось под действием 1 мг фермента в мин [8].
Активность супероксиддисмутазы (СОД) выявляли по ингибированию ее молекулами фотохимического восстановления нитросинего тетразолия (НСТ). Единица активности соответствовала количеству фермента, способному подавить реакцию восстановления НСТ на 50 % [1].
Для определения содержания вторичных продуктов ПОЛ мы применяли стандартный подход, основанный на их взаимодействии с 2-тиобарбитуровой кислотой (ТБК) с образованием окрашенного продукта – малонового диальдегида (МДА) [9].
Элементный анализ почвенных и растительных образцов проводили в Центре коллективного пользования научным оборудованием Воронежского государственного университета с помощью растрового электронного микроскопа Jeol JSM-680LV. Образцы предварительно высушивали, измельчали и помещали под гидравлический пресс для получения однородных по составу поверхностей.
Статистическую обработку результатов проводили при уровне значимости 5 % с использованием t-критерия Стьюдента.
Результаты исследования и их обсуждение
Одним из важных показателей эффективности ферментативного звена АОС является сбалансированность активности СОД, каталазы и пероксидазы. Подавление активности одного из ферментов антиоксидантной системы может привести к избыточному накоплению АФК и деструкции клеток.
Супероксиддисмутаза катализирует превращение супероксидных радикалов в Н2О2, играет ключевую роль в утилизации свободных радикалов и предотвращении оксидативного повреждения клетки. Принято считать, что основным регулятором активности СОД в клетке является уровень О2•–, который выступает по отношению к ферменту в качестве индуцирующего фактора, причем индукция СОД при повышении генерации О2•– связана с усилением синтеза фермента в клетке de novo [6].
Из рис. 1 видно, что общая СОД-активность при произрастании на черноземе и в условиях искусственно созданной модели альпийской горки не изменялась в листьях двух из исследуемых нами видов: Бурачок Гмелина (Alyssum gmelinii) и Лен многолетний (Linum perenne). Экстракты Шлемника приземистого (Scutellaria supina) при его произрастании на черноземе проявляли более высокую супероксиддисмутазную активность.
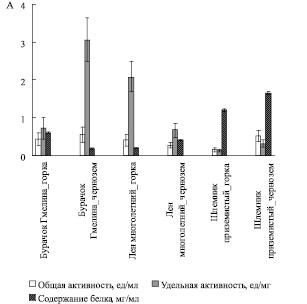
Рис. 1. Активность супероксиддисмутазы (А)
Для более корректной трактовки полученных результатов мы определили содержание белка в растительных пробах, чтобы вычислить удельную активность ферментов антиоксидантной защиты. Удельная активность супероксиддисмутазы в листьях видов Бурачок Гмелина и Шлемник приземистый, произрастающих на черноземе, оказалась выше, чем в условиях горки. Для льна многолетнего – напротив, значения удельной СОД активности на черноземе были ниже.
Вторым звеном защиты от АФК служат ферменты, удаляющие Н2О2: каталаза и пероксидазы. Эти энзимы, используя в качестве донора электронов H2O2 в случае каталазы или различные органические соединения в случае пероксидазы, катализируют двухэлектронное восстановление Н2О2 до H2O [5].
Из рис. 2 следует, что при произрастании в условиях горки общая каталазная активность увеличивалась в листьях шлемника приземистого (Scutellaria supina) и не изменялась у двух других исследуемых нами видов. Удельная активность каталазы увеличивалась в условиях горки в листьях шлемника приземистого (Scutellaria supina) и льна многолетнего (Linum perenne), в листьях же бурачка Гмелина (Alyssum gmelinii), напротив, наблюдалось снижение удельной каталазной активности.
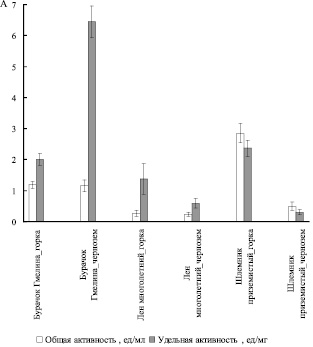
Рис. 2. Активность каталазы (А)
Из рис. 3 видно, что общая и удельная активность пероксидазы при произрастании видов Шлемник приземистый и Бурачок Гмелина в условиях горки снижалась; в отличие от них в экстрактах льна многолетнего общая активность пероксидазы в условиях горки не изменялась, а удельная – увеличивалась.
Изложенные выше результаты позволяют констатировать, что при произрастании в условиях горки ферменты антиоксидантной защиты в листьях шлемника приземистого и бурачка Гмелина в основном менее активны, чем при произрастании этих видов на черноземе. У льна многолетнего, напротив, при росте в условиях горки наблюдалось увеличение значений удельной активности каталазы, пероксидазы и супероксиддисмутазы при отсутствии статистически достоверных изменений в их общей активности.
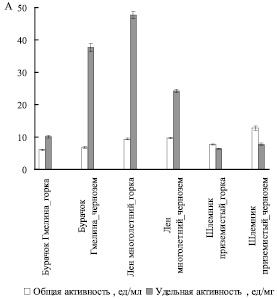
Рис. 3. Активность пероксидазы (А)
Исходя из данных по активности ферментов антиоксидантной системы, сложно однозначно судить об оксидативном статусе растения, эти сведения необходимо сопоставить с материалами о содержании активных форм кислорода или промежуточных продуктов их распада в исследуемом объекте. К вторичным продуктам ПОЛ относят такие кислородосодержащие соединения, как спирты, альдегиды и диальдегиды, лактоны, эпоксиды и другие вещества, которые образуются в результате распада липидных пероксидов. В биологических системах эти продукты находятся обычно в достаточно высоких стационарных концентрациях и принимают участие в различных биохимических процессах.
В следующей серии экспериментов мы определили концентрацию малонового диальдегида (вторичного продукта ПОЛ) в экстрактах листьев исследуемых нами растений.
Из рис. 4 отчетливо видно, что концентрация МДА снижалась при произрастании в условиях искусственно созданной горки только в листьях Шлемника приземистого (Scutellaria supina), у двух других видов статистически достоверных изменений не наблюдалось.
Интересно, что при произрастании как на горке, так и на черноземе в листьях льна многолетнего концентрация МДА была одинакова при том, что удельная активность всех трех исследуемых нами ферментов антиоксидантной защиты в условиях горки выше. Это может косвенно свидетельствовать о том, что растению на горке необходимо использовать больший внутренний ресурс для поддержания той же концентрации активных форм кислорода, чем при росте на черноземе.
У Бурачка Гмелина в условиях роста на обоих видах почв при одинаковой концентрации МДА удельная активность всех трех ферментов на горке ниже, а общая либо не изменялась, либо также была ниже. Этот факт позволяет предположить, что условия искусственно созданной альпийской горки являются более благоприятными для произрастания данного вида, чем чернозем.
Аналогичное предположение справедливо и для Шлемника приземистого: при произрастании на горке, наряду с понижением концентрации МДА, мы наблюдали снижение активности СОД и пероксидазы.
В дополнение к результатам предыдущих экспериментов мы осуществили элементный анализ растительных и почвенных образцов, так как состав почвы может в значительной степени оказывать влияние на оксидативный статус растения [4]. Установлено, что содержание кальция ( %) в пробах почвы с горки значительно (~ 3,3 раза) превышает его количество в черноземе. Интересно, что в условиях произрастания на горке (по сравнению с условиями чернозема) количество кальция ( %) выше в листьях Льна многолетнего и Шлемника приземистого, но ниже у Бурачка Гмелина (рис. 5).
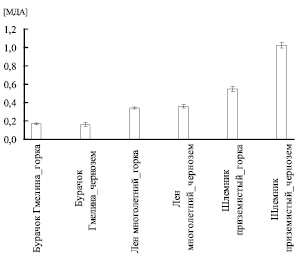
Рис. 4. Количество малонового диальдегида [МДА] в экстрактах листьев, моль
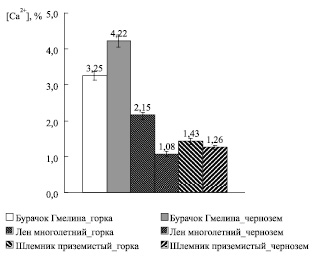
Рис. 5. Количество кальция ( %) в листьях растений
При действии на растения неблагоприятных и повреждающих факторов среды происходят изменения содержания и соотношения пигментов, определяющих цвет листьев. Имея сведения о содержании хлорофилла, можно оценить эффективность использования растениями фотосинтетически активной радиации. Характерные изменения окраски листьев, такие как хлороз, пожелтение, некроз, побурение, являются симптомами неблагополучия растений. Старение листьев сопровождается сохранением значительной части каротиноидов или активацией их синтеза, что происходит на фоне разрушения хлорофилла [2].
Из рис. 6 следует, что при произрастании на горке в листьях шлемника и бурачка концентрация каротиноидов была ниже, а отношение суммарной концентрации хлорофиллов к концентрации каротиноидов выше, чем при их росте на черноземе, что может свидетельствовать о более благополучных условиях произрастания на горке для этих видов.
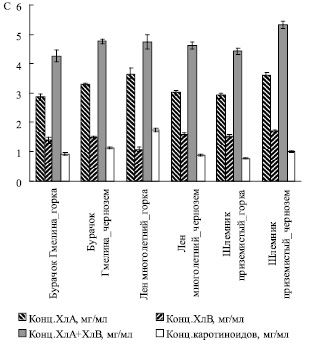
Рис. 6. Содержание хлорофиллов и каротиноидов (С) в листьях растений
Заключение
Исходя из результатов всех проведенных нами экспериментов, можно заключить, что на искусственно созданной альпийской горке условия произрастания для видов Бурачок Гмелина (Alyssum gmelinii) и Шлемник приземистый (Scutellaria supina) более благоприятны, чем на черноземе, тогда как для льна многолетнего (Linum perenne) по использованным нами биохимическим показателям не удалось установить более предпочтительный для него вид почвы. В дальнейшем нам представляется необходимым расширить перечень биохимических показателей и ассортимент экспериментальных объектов исследования.
Работа выполнена при поддержке Минобрнауки России в рамках государственного задания ВУЗам в сфере научной деятельности на 2014 – 2016 годы. Проект № 1035.
Рецензенты:
Епринцев А.Т., д.б.н., профессор, зав. кафедрой биохимии и физиологии клетки Воронежского государственного университета, г. Воронеж;
Девятова Т.А., д.б.н., профессор, зав. кафедрой экологии и земельных ресурсов Воронежского государственного университета, г. Воронеж.
Работа поступила в редакцию 04.06.2014.
Библиографическая ссылка
Холявка М.Г., Карпова С.С., Калаев В.Н., Лепешкина Л.А., Агапов Б.Л., Артюхов В.Г. ОЦЕНКА ОКСИДАТИВНОГО СТАТУСА РАСТЕНИЙ, ПРОИЗРАСТАЮЩИХ В РАЗЛИЧНЫХ УСЛОВИЯХ // Фундаментальные исследования. – 2014. – № 8-4. – С. 891-897;URL: https://fundamental-research.ru/ru/article/view?id=34690 (дата обращения: 19.04.2024).