



Показано, что пребывание в среде с пониженной гравитационной нагрузкой сопровождается снижением тонуса и силы сокращения мышц [4, 32, 33, 35], суставной чувствительности [16], нарушением координации движений [2, 3] и рефлекторных механизмов [38]. Общепризнано, что главным фактором, ответственным за все эти изменения, является внезапное устранение проприоцептивной информации от мышц и сухожилий в ответ на отсутствие механической нагрузки, и, соответственно, в этой связи антигравитационные или постуральные мышцы являются основной мишенью для действия функциональной разгрузки.
Космические полеты сопровождаются большой потерей массы мышц-разгибателей, составленных преимущественно из медленно сокращающихся волокон, умеренной потерей в мышцах-разгибателях, составленных преимущественно из быстросокращающихся волокон и наименьшими потерями в мышцах-сгибателях, составленных преимущественно из быстро сокращающихся волокон [27]. Эти различия базируются на распределении типов волокон мышцы. В исследованиях [3, 46] показано, что невесомость, моделируемая водным погружением, изменяет порядок рекрутирования двигательных единиц при выполнении изометрических произвольных сокращений. Гравитационная нагрузка, по-видимому, необходима для сохранения размера (массы) и силы скелетных мышц у человека [30, 34, 40]. В исследованиях, моделирующих условия микрогравитации [7, 30, 32, 33], показано, что физическая тренировка уменьшает, но не предотвращает полностью потерю массы и сократительных свойств мышц. Наибольшему влиянию микрогравитации подвержены антигравитационные мышцы-разгибатели бедра и особенно стопы [2, 7], вероятно, из-за большей их механической нагрузки в нормальных гравитационных условиях. Непропорционально большая потеря силы мышцы по сравнению с ее размером [10, 30] прямо указывает на существование других факторов, которые вносят существенный вклад в ослабление мышцы кроме общепризнанной атрофии.
Внутренняя архитектура мышцы - важный детерминант функциональных характеристик мышцы (отношение скорость - сила, длина-сила и максимальная изометрическая сила (например, [22, 41]).
Трехглавая мышца голени - это основной синергист подошвенного сгибания стопы, которая обладает разными архитектурными свойствами, такими, как длина мышцы, длина мышечного пучка/волокна и угол наклона [13, 19]. Кроме того, икроножные мышцы - двусуставные, тогда как камбаловидная - односуставная. Следовательно, взаимоотношение между суставными углами (коленным и голеностопным), длиной мышцы/пучка и углами наклона является высокоспецифичными для каждой мышцы. Информация относительно изменений внутренней архитектуры мышц, связанной с позицией суставного угла, является важной при изучении функций мышцы.
Цель настоящего исследования - сравнить изменения в архитектуре разных головок, составляющих трехглавую мышцу голени [медиальную (МИМ) и латеральную (ЛИМ), икроножные мышцы и камбаловидную (КМ) мышцу] у человека и обсудить функциональные значения этих изменений после 7-суточной «сухой» водной иммерсии с использованием продолжительной функциональной электромиостимуляционной тренировки.
Методы и материал
Испытуемые и «сухая» водная иммерсия
В исследовании приняла участие группа из шести практически здоровых мужчин-добровольцев в возрасте 22,8 ± 0,8 лет, ростом и массой 1,84 ± 0,1 м и 79,3 ± 4,2 кг, соответственно. Все испытуемые клинически были оценены как здоровые и находились в хорошем физическом состоянии, не употребляли медикаментозных средств и были некурящими. Отбор испытуемых базировался на анализе истории болезни, биохимического анализа крови, анализа электрокардиограммы в покое и при выполнении велоэргометрического нагрузочного теста.
Экспериментальные процедуры были выполнены в соответствии с Хельсинской декларацией, и испытуемые после полной информации о процедурах и задачах настоящего исследования дали письменное согласие на участие в нем. Программа исследований была одобрена комиссией по биомедицинской этике при ГНЦ РФ - ИМБП РАН.
Для моделирования воздействия функциональной разгрузки мышечного аппарата использовали модель «сухой» водной иммерсии [5]. Испытуемого помещали в горизонтальном положении в специальную ванну с высокоэластической тканевой пленкой, изолирующей поверхность тела от жидкости - воды. Температура воды в ванне была постоянной на уровне 33,4 ºC и автоматически поддерживалась на этом уровне в течение всего эксперимента. Испытуемый постоянно оставался в горизонтальном положении, включая выделительные функции и прием пищи. Во время экспозиции испытуемый постоянно находился под медицинским наблюдением на протяжении 24 час. Обслуживающий медицинский персонал постоянно присутствовал при транспортировании испытуемого, при выполнении личной гигиены, включая туалет и прием пищи, медицинском обслуживании в пределах ограничений протокола.
Сократительные свойства трехглавой мышцы голени (ТМГ) исследовали дважды - до и после иммерсионного воздействия, и протокол исследований был идентичен.
Измерение силы
Измерения были выполнены на правой, ведущей у всех испытуемых, конечности. Испытуемый удобно располагался в положении «стоя на коленях» на специальном кресле изокинетического динамометра (модель «Biodex», USA), и стопа его правой конечности относительно жестко фиксировалась к измерительной платформе динамометра. При этом стопу испытуемого крепили таким образом, чтобы ось голеностопного сустава совпадала с осью вращения педали динамометра.
В процессе измерения максимального суставного момента каждого испытуемого тщательно инструктировали, как выполнять подошвенное сгибание стопы. По команде «сократить максимально сильно» испытуемый выполнял три ритмических изометрических подошвенных сгибаний из положения голеностопного сустава - 0º (нейтральное анатомическое положение голеностопного сустава) и коленного сустава - 90º.
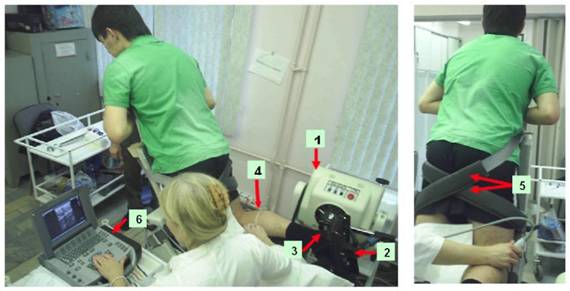
Рис. 1. Экспериментальная установка
Позиция испытуемого при тестировании функций на изокинетическом динамометре и ультразвуковых исследований МИМ и КМ. При тестировании функций голеностопный и коленный суставы находятся в позиции 90º (нейтральное анатомическое положение) и 60º, соответственно; 1 - регистрирующее устройство изокинетического динамометра; 2 - платформа для фиксации стопы испытуемого; 3 - ось, соединяющая регистрирующее устройства динамометра с силовой платформой; 4 - маркер (стрелка) между кожей и ультразвуковым датчиком, служащий ориентиром движения датчика во время исследования; 5 - ремни для фиксации бедра; 6 - ультразвуковая система.
Положение ультразвукового датчика при исследовании МИМ (левая панель) и ЛИМ (правая панель)
Функциональная электрическая стимуляция
Принцип тренировки
Функциональная электрическая стимуляция (ФЭС) была применена к мышцам нижних конечностей и обеспечивалась двумя электростимуляторами «СТИМУЛ НЧ-01» (РОССИЯ), генерирующими двухполярные симметричные прямоугольные электрические импульсы длительностью 1 мс, частотой 25 Гц и амплитудой от 0 до 45 В. Техническое оборудование для ФЭС-тренировки (ФЭСТ) мышц состояло из специальных брюк с клапанами для электродов, двух шестиканальных электростимуляторов, соединенных между собой кабелем синхронизации, и «сухих» электродов стимуляции (фирма «Axelgaard», USA).
После подготовки кожи два слегка увлажненных «сухих» электрода устанавливали на m. quadriceps femoris, m. hamstrings, m. tibialis anterior, m. peroneal и m. triceps surae. Синхронная стимуляция всех мышц предотвращала нежелательные движения конечностей.
Испытуемого тщательно инструктировали о порядке пользования электростимуляторами. ФЭСТ мышц передней и задней поверхности бедра и голени каждой конечности осуществлялась непосредственно при экспозиции испытуемого в ванне на протяжении 6 суток по 3 часа в день. Длительность сокращения мышц при ФЭС составляла 1 с и интервал отдыха между сокращениями - 2 с.
Ультразвуковое сканирование
Угловая позиция сустава и измерение суставного момента
Для определения суставных моментов при выполнении произвольных изометрических сокращений мышц-разгибателей стопы использовали изокинетический динамометр (модель «Biodex», USA). Все измерения были выполнены на правой конечности испытуемого (рис. 1). Стопа правой исследуемой конечности относительно жестко крепилась к специальной платформе измеряемого устройства изокинетического динамометра так, чтобы ось вращения измеряемого устройства динамометра совмещалась с осью вращения голеностопного сустава (центр боковой лодыжки). Коленный сустав находился в позиции 90 °. Угол голеностопного сустава последовательно был установлен в позицию -15 ° подошвенного сгибания, 0 ° (нейтральная анатомическая позиция), +15 и +30 ° подошвенного разгибания. В каждой позиции испытуемого просили расслабить ТМГ (пассивное состояние), и установленный пассивный суставной угол регистрировался программным обеспечением компьютера изокинетического динамометра. Затем в пассивном состоянии голеностопный сустав устанавливали в нейтральную анатомическую позицию (0°), и испытуемый выполнял 3-5 субмаксимальных сокращений для ознакомления с экспериментальной процедурой. После этого испытуемого просили по команде «сократить максимально сильно» выполнить максимальное произвольное изометрическое разгибание стопы (активное состояние) и регистрировали суставной момент (изометрическое максимальное произвольное сокращение - МПС). Затем просили выполнить сокращение с усилием 50 % от МПС (активный режим) при нейтральной позиции голеностопного сустава (0 °) и удерживать сокращение на протяжении 2-3 с. Каждый испытуемый выполнял три попытки (с интервалом между сокращением в 1 мин), и лучшую попытку использовали для дальнейшего анализа. Кроме того, развиваемые мышечные усилия испытуемый визуально контролировал по экрану монитора.
Измерение длины, угла перистости волокна и толщины мышцы
Длина и угол перистости волокон ТМГ у человека были измерены в условиях in vivo при помощи ультразвукового сканирования во время отдыха (пассивное состояние) и при развитии усилия, составляющего 50 % МПС (активное состояние). Для исследования мышечной архитектуры в реальном времени использовался B-режим изображения ультразвуковой диагностической портативной системы (модель «SonoSite MicroMaxx», USA) с электронным линейным датчиком 7,5 МГц, толщиной 10 мм и длиной сканирующей поверхности 60 мм, чтобы получить сагиттальное изображение МИМ, ЛИМ и КМ в покое и при усилии 50 % от МПС при нейтральной анатомической позиции голеностопного сустава (0°). Для лучшего акустического сцепления сканирующую поверхность датчика и мышцы покрывали специальным гелем и датчик ориентировали по среднесагиттальной оси мышцы. Качество ультразвукового изображения мышечного пучка достигалось регулировкой глубины и яркости ультразвукового сигнала. В покое и во время сокращения датчик удерживался на брюшке мышцы. Во время сканирования, чтобы не нарушить мышечную структуру, применяли минимальное давление датчика на кожные покровы мышцы. Во время измерений в условиях покоя испытуемых инструктировали - «полностью расслабить мышцу конечности». Визуализация мышц осуществлялась с использованием панорамного и фрагментарного изображения мышцы в продольном или поперечном сечении (области наибольшей анатомической площади поперечного сечения). Для получения фрагментарного изображения продольного сечения мышцы датчик фиксировался перпендикулярно к поверхности мышцы и продольно в области наибольшей анатомической площади поперечного сечения мышцы с «захватом» анэхогенной линии маркера, расположенного по окружности голени. Маркер представлял собой медную проволоку (∅ 1,5 мм; рис. 1, левая панель) в мягкой пластиковой оболочке и располагался по окружности голени на уровне (см. ниже), соответствующем наибольшей анатомической площади поперечного сечения мышцы [21]. Для получения фрагментарного изображения поперечного сечения мышцы датчик располагался перпендикулярно к поверхности мышцы вдоль поперечной оси мышцы. Для панорамного изображения продольного сечения мышцы датчик фиксировался на маркере и плавно перемещался по продольной оси в направлении от проксимального к дистальному концу мышцы, а для получения фрагментарного изображения поперечного сечения мышцы датчик перемещался по окружности задней поверхности голени в поперечном направлении вдоль маркера.
В позиции пассивного и активного режимов были получены продольные ультразвуковые изображения МИМ, ЛИМ и КМ на уровнях, соответствующих 30 % (МИМ и ЛИМ) и 50 % (КМ) расстояния между подколенной складкой и центром боковой лодыжки. На этих уровнях помещался маркер, являющийся ориентиром того, что датчик во время измерений не смещался. Ультразвуковые изображения записывались на жесткий диск ультразвуковой системы (модель «SonoSite MicroMaxx», USA) для дальнейшего анализа.
Длина волокна (Lв) мышцы (рис. 2) определялась как расстояние между местом прикрепления волокна у поверхностного апоневроза до места вхождения его в глубокие слои апоневроза мышцы [20]. Визуализируя по ультрасонограмме пучки по их длине от поверхностного к глубоким слоям апоневроза, можно проверить правильность сканирования пучка [44], в противном случае длина пучка может быть оценена слишком завышенной, а угол пучка недооценен [21].
Угол перистости (Θв) волокон (рис. 2) определялся как угол, создаваемый линией, проведенной от точки прикрепления волокна у поверхностного слоя апоневроза к месту прикрепления волокна к глубокому слою апоневроза [43].
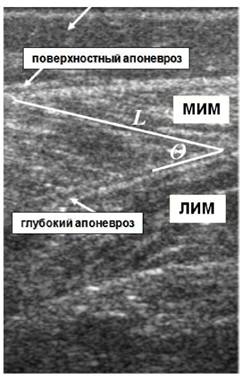
Рис. 2. Ультразвуковые изображения продольного сечения медиальной икроножной мышцы (МИМ), латеральной икроножной мышцы (ЛИМ) и камбаловидной мышцы (КМ). Представлен угол (Θ) перистости и длина (L) волокна между глубоким и поверхностным апоневрозами
У каждого испытуемого анализировались параметры не менее трех мышечных пучков, а затем полученные данные усреднялись и анализировались. Коэффициент вариации трех измерений находился в диапазоне 0-2 %.
Толщина мышцы определялась по уравнению:
толщина мускула=Lxsin α
где, L и α - каждой мышцы определялись ультразвуком
Показатель внутреннего укорочения мышечного волокна (∆Lмышца) [29] при сокращении мышцы определялся по разнице между L и cos Θ перистости волокна при активном состоянии мышцы по сравнению с ее пассивным состоянием.
Статистика
При обработке полученных результатов исследования применяли общепринятые статистические методы - рассчитывали среднюю и стандартную ошибку средней (М ± m). Достоверность различий оценивали с помощью параметрического t-критерия Стьюдента и уровень достоверности р < 0,05 признавался как существенный.
Ультразвуковые изображения обрабатывались с использованием пакета программ «Dr. ReallyVision» (ООО «Альянс-Холдинг», Россия).
Результаты
Изменение максимальной мышечной силы
Максимальный суставной момент, развиваемый мышцами-разгибателями стопы после иммерсии с применением ФЭСТ, увеличился в среднем на 14.4 % (146.9 ± 8.3 против 168,1 ± 4,3 Н) и незначительно (~9,6 %) уменьшился (155 против 140 Н) у одного испытуемого (рис. 3)
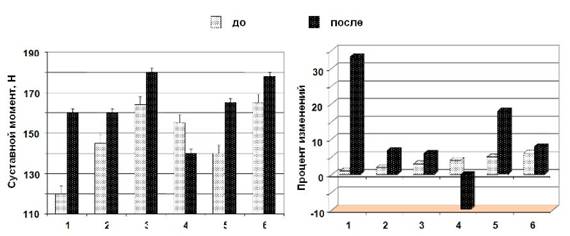
Рис. 3. Максимальный суставной момент (панель слева) и процент изменений (панель справа), развиваемый трехглавой мышцей голени в результате ФЭСТ в условиях «сухой» водной иммерсии
Архитектурная характеристик в условиях покоя
Толщина МИМ, ЛИМ и КМ в условиях покоя (положение голеностопа -15 °) значительно не изменялась (~18, 16 и 15 мм, соответственно) в ответ на изменения L мышцы при изменении угла в голеностопном суставе (рис. 4).

Рис. 4. Изменение толщины МИМ, ЛИМ и КМ как функция угла в голеностопном суставе
Θв и Lв во всех трех мышцах в покое были зависимы от угла голеностопного сустава (таблица 1). Во всех мышцах, когда угол в голеностопном суставе изменялся от -15 ° до +30°, то Θв увеличился в МИМ от 31 ± 2,8 до 49 ± 1,7 ° (58 %, p < 0·01), в ЛИМ от 20 ± 2,1 до 28,5 ± 1,6° (43 %, p < 0·05) и в КМ от 22,8 ± 1,4 до 34 ± 2,2° (49 %, p < 0·01) (рис, 5),
Во всех трех мышцах при увеличении угла в голеностопном суставе от -15° до +30°
Lв уменьшилась: в МИМ от 36 ± 1.2 до 27 ± 2,1 мм (25 %, p < 0·01), в ЛИМ от 46,8 ± 0,6 до 31,2 ± 1,9 мм (33 %, p < 0·01) и в КМ от 39,2 ± 1,2 до 28,2 ± 2,0 мм (28 %, p < 0·01) (таблица 1; рис, 5).
Архитектурные характеристики после иммерсии
После ИМ с применением НЭМС Н МИМ, ЛИМ и КМ при изменении угла в голеностопном суставе от -15 до +30 ° была снижена по сравнению с исходной (рис, 4).
Во всех трех мышцах после иммерсии с ФЭСТ Θв и Lв были зависимы от угла голеностопного сустава (таблица 1; рис, 5). Во всех трех мышцах, когда угол в голеностопном суставе увеличивался от -15° до +30°, Θв увеличился - в МИМ от 26 ± 2,8 до 36 ± 2,8° (38 %, p < 0,05), в ЛИМ от 15 ± 1,4 до 20,2 ± 1,0° (35 %, p < 0,05) и в КМ от 18,5 ± 1,6 до 24,8 ± 1,7° (34 %, p < 0,01) (таблица 1; рис, 5).
Во всех трех мышцах при увеличении угла в голеностопном суставе от -15° до +30° Lв уменьшилась: в МИМ от 30,2 ± 1,7 до 25,5 ± 1,9 мм (16 %), в ЛИМ от 40,2 ± 0,6 до 25,2 ± 2,9 мм (37 %, p < 0,01) и в КМ от 29,2 ± 2,9 до 22,2 ± 2,1 мм (24 %, p < 0,05) (таблица 1; рис, 5).
Угол перистости и длина волокон в МИМ, ЛИМ и КМ
|
мышца / угловая позиция |
до |
после |
∆, % |
|
длина волокна, мм |
|
|
|
|
МИМ |
|
|
|
|
-15 |
35 ± 1 |
30 ± 2 |
15 |
|
0 |
32 ± 2 |
28 ± 1 |
12 |
|
15 |
30 ± 2 |
26 ± 3 |
13 |
|
30 |
27 ± 2 |
25 ± 2 |
5 |
|
ЛИМ |
|
|
|
|
-15 |
47 ± 1 |
40 ± 1 |
14 |
|
0 |
36 ± 2 |
31 ± 2 |
13 |
|
15 |
32 ± 2 |
25 ± 2 |
22 |
|
30 |
31 ± 2 |
25 ± 3 |
19 |
|
КМ |
|
|
|
|
-15 |
39 ± 1 |
29 ± 3 |
26 |
|
0 |
36 ± 3 |
32 ± 2 |
13 |
|
15 |
34 ± 1 |
28 ± 2 |
17 |
|
30 |
28 ± 2 |
22 ± 2 |
21 |
Угол перистости, °
|
МИМ |
|
|
|
|
-15 |
31 ± 3 |
26 ± 3 |
15 |
|
0 |
37 ± 3 |
29 ± 2 |
22 |
|
15 |
42 ± 5 |
32 ± 4 |
25 |
|
30 |
49 ± 2 |
36 ± 3 |
26 |
|
ЛИМ |
|
|
|
|
-15 |
20 ± 2 |
15 ± 1 |
24 |
|
0 |
22 ± 2 |
18 ± 1 |
20 |
|
15 |
22 ± 1 |
18 ± 2 |
19 |
|
30 |
28 ± 2 |
20 ± 1 |
29 |
|
КМ |
|
|
|
|
-15 |
23 ± 1 |
18 ± 2 |
19 |
|
0 |
28 ± 2 |
24 ± 3 |
16 |
|
15 |
30 ± 2 |
22 ± 3 |
26 |
|
30 |
34 ± 2 |
25 ± 27 |
27 |
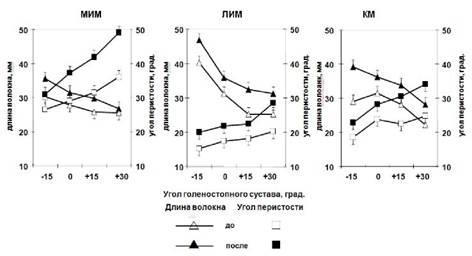
Рис. 5. Архитектура трехглавой мышцы голени. Изменение длины и угла перистости волокон МИМ, ЛИМ и КМ как функция угла в голеностопном суставе.
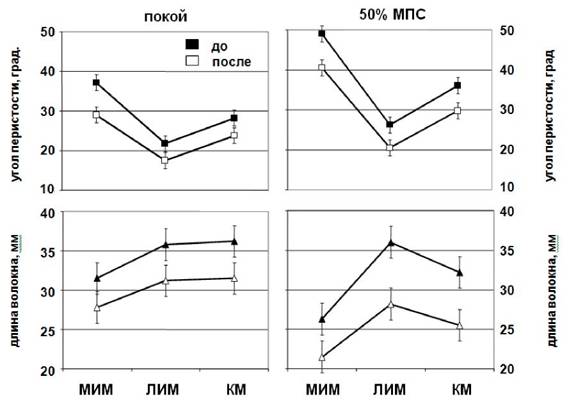
Рис. 6. Архитектурная характеристика во время развития градуируемой силы
Изменения длины волокон МИМ, ЛИМ и КМ в покое и при усилии 50 % МПС при нейтральном анатомическом положении голеностопного сустава (0º)
Архитектурные характеристики во время развития градуируемой силы
Изменение Θв и Lв как функция угла голеностопного сустава 90° от условий покоя к усилию 50 % от МПС представлено на рис. 6.
Θв и Lв волокон уменьшались как функция интенсивности сокращения во всех трех мышцах, Θв в МИМ, ЛИМ и КМ постепенно уменьшался с 49 ± 1,4 до 40,5 ± 2,4 ° (17 %, p < 0,01), от 26,2 ± 1,8 до 20,5 ± 1,9 ° (22 %, p < 0,05) и от 36 ± 5,3 до 29,8 ± 3,9 ° (17 %), соответственно (рис. 6). Lв в МИМ, ЛИМ и КМ постепенно уменьшилась с 26,3 ± 2,6 до 21,5 ± 1,9 мм (18 %), от 36 ± 2,9 до 28,2 ± 2,2 мм (22 %, p < 0,05) и от 32,2 ± 2,2 до 25,5 ± 2,1 мм (21 %, p < 0,05), соответственно (рис. 6).
Сравнение между фактическими и архитектурными изменениями после иммерсии Θв во время 50 % МПС.
Оцененный после иммерсии с ФЭСТ Θв во время 50 % МПС отличался: 8,9 (17,3 %, p < 0,05), 5,7 (21,8 %, p < 0,05), и 6,2° (17,2 %, p < 0,05), от соответствующих фактических Θ перистости в МИМ, ЛИМ и КМ соответственно (рис. 6).
В МИМ величины Θв были выше, чем соответствующие фактические величины при сокращении с интенсивностью 50 % МПС, но в ЛИМ и КМ величины были систематически ниже, чем соответствующие фактические величины.
Lв во время 50 % МПС.
Величина Lв во время 50 % МПС отличались на 4,8 мм (18,2 %, p < 0,01), 7,8 мм (21,7 %, p < 0,01) и 6,7 мм (20,8 %, p < 0,01) от соответствующих фактических величин МИМ, ЛИМ и КМ соответственно (рис. 6).
Внутреннее укорочение волокон
Более короткие Lв и более крутые Θв при активном состоянии по сравнению с пассивным показывают степень внутреннего укорочения волокон во время сокращения.
До иммерсии ∆Lмышцы МИМ составила 7,9 мм, а после иммерсии с ФЭСТ уменьшилась и составила 7,8 мм, в КМ - уменьшилась от 5,9 против 5,6 мм. Существенное увеличение ∆Lмышцы обнаружено в ЛИМ - от 0,9 до 3,3 мм.
Обсуждение
Настоящее исследование впервые описывает архитектуру разных головок ТМГ (МИМ, ЛИМ И КМ) у человека в условиях in vivo в покое и во время градуируемого изометрического подошвенного сгибания (50 % МПС).
Результаты, полученные в условиях in vivo, показывают, что архитектура МИМ, ЛИМ и КМ у человека существенно изменяется и как функция угла голеностопного сустава в покое и как функция силы, развиваемая во время изометрических сокращениях при установленном суставном угле. В покое при пассивном изменении угла в голеностопном суставе от -15 до +30° угол перистости волокон МИМ увеличивается от 31 до 49°, ЛИМ - от 20 до 28,5° и КМ - от 22,8 до 34°, а длина волокон МИМ уменьшается от 35,5 до 26,8 мм, ЛИМ - от 46,8 до 31,2 мм, и КМ - от 39,2 до 28,2 мм. Эти результаты показывают, что длина и угол перистости волокон ТМГ у человека не остаются постоянными при изменяющейся длине мышцы [26]. Уменьшение длины волокон мышц при пассивном изменении угла в голеностопном суставе от -15 до +30° предполагает, что волокна мышцы при увеличении угла в суставе прогрессивно «ослабевают» [8]. Ichinose et al. [26], используя ультразвуковой метод у человека, на примере m. vastus lateralis показали, что в условиях покоя «ослабление» волокон мышцы является функцией угла сустава. Авторы отметили, что, когда угол в коленном суставе составляет 180°, т.е. отмечается полное разгибание колена, волокна мышцы значительно ослаблены, поскольку они уменьшаются примерно до 35 % своей длины, составляя величину сокращения лишь 10 % МПС. В данном исследовании уменьшение длины мышечных волокон (ср. рис. 5 и таблицу. 1) при пассивном изменении угла в голеностопном суставе от -15 до +30 ° предполагает, что волокна мышцы стали более «слабыми».
В исследовании показано, что переход от условий покоя к усилию 50 % МПС угол перистости волокон МИМ увеличивается от 37,2 до 49,0 °, а длина волокна уменьшается
с 31,5 до 26,9 мм; угол перистости волокон ЛИМ увеличивается от 21,8 до 26,2 °, а длина волокна немного увеличивается с 35,8 до 36,0 мм в КМ угол перистости увеличивается от 28,2 до 36,0 мм, а длина волокна уменьшается с 36,2 до 32,2 мм без существенного изменения в расстоянии между апоневрозами. Полученные данные хорошо согласуются с предсказаниями Gans и Bock [22]), согласно которым «...увеличение перистости подготавливает, чтобы волокна дали компенсацию за изменение в угле волокна во время сокращения...».
Целью настоящего исследования было попытаться объяснять влияние хронической разгрузки на механические свойства ТМГ у человека и оценить вклад профилактического влияния, оказываемое ФЭСТ, выполненной во время разгрузки, на сократительные свойства мышцы. Наши результаты показывают, что дефицит механической нагрузки, или иначе функциональная разгрузка мышечного аппарата, вызывает снижение структурных и увеличение сократительных свойств ТМГ у человека, и, хотя используемый режим тренировки/упражнений (ФЭСТ) действительно уменьшал отрицательное влияние функциональной разгрузки, но полностью их не предотвратил. Тем не менее настоящее исследование можно считать уникальным в терминах продолжительности разгрузки с использованием ФЭСТ.
Многие исследования показывают, что условия невесомости (отсутствие весовой нагрузки) вызывают изменения сократительных функций мышц [2, 6, 18, 28, 30-36, 39]. Более того, механическая разгрузка, вызванная условиями «сухой» водной иммерсии, сопровождается изменением порядка рекрутирования двигательных единиц (ДЕ) во время выполнения произвольных изометрических сокращений [3, 46]. Порядок рекрутирования ДЕ полностью не установлен, будучи переменный при различных условиях, но один из основных факторов, управляющий порядком рекрутирования ДЕ и являющийся наиболее важным, - это вклад афферентного входа от проприорецепторов к мотонейронам, особенно при произвольном сокращении мышцы, поскольку невесомость освобождает мышцы от ее весовой задачи, и это должно уменьшить входы (вклад) проприорецепторов от веретен и сухожилия мышцы.
Основной результат настоящей работы в том, что после иммерсии с применением ФЭСТ величина изометрического максимального суставного момента, развиваемого мышцами-разгибателями стопы, увеличилась (~14 %), тогда как предыдущие исследования показали, что в отсутствие профилактических мероприятий существенно уменьшается МПС - более чем на 50 % [2, 32, 33, 35] и Pо - больее чем на 30 % [32, 33, 35]. Эффективность ФЭСТ, как было показана ранее [31, 37, 42], - это значительное увеличение функциональных свойств мышц в относительно короткий срок. В настоящем исследовании незначительное увеличение силы сокращения мышц у испытуемых, возможно, определяется тем, что в среднем интенсивность применяемых стимуляционных импульсов во время тренировки была чрезвычайно недостаточной для активации ДЕ, и в частности малых ДЕ (см. рис. 7, испытуемый 4).
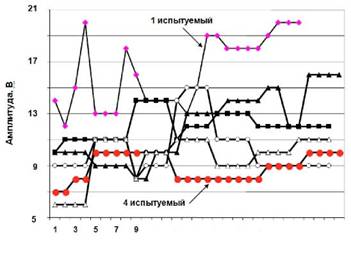
Рис. 7. Динамика изменения амплитуды стимуляционного импульса на протяжении ФЭСТ
Известно, что когда сила сокращения мышцы прогрессивно увеличивается, то малые мотонейроны, иннервирующие мышцы, быстрее активируются, чем большие клетки [17, 25]. Малые ДЕ состоят из медленносокращающихся волокон типа I, а большие - из быстросокращающихся типа II. При субмаксимальных произвольных сокращениях волокна ДЕ типа I активируются синоптическим потоком, оканчивающимся на моторном нейроне. Ситуация полностью отличается в сокращениях, вызванных ФЭС, поскольку в этом случае волокна ДЕ активируются электрическим током, который применен внеклеточно к окончаниям нерва, и большие мотонейроны с более низким аксональным входным сопротивлением становятся более легковозбудимыми [15, 45]. Фактически, когда стимул применен снаружи клетки, электрический ток, сначала должен поступить через мембрану прежде, чем деполяризуется клетка, но внеклеточная среда шунтирует ток и меньшие ДЕ не будут активированы во время субмаксимальной ФЭС из-за более их высокого аксонального входного сопротивления. Поэтому меньшие ДЕ не тренируются при субмаксимальной ФЭС. Однако, когда используется электрическое возбуждение высокой интенсивности, большой силой стимулирующего импульса, ФЭС будет более эффективным упражнением [8].
Увеличение максимального суставного момента сопровождалось изменениями внутренней архитектуры МИМ, ЛИМ и КМ, которая была частично предотвращена ФЭСТ. После иммерсии с применением ФЭСТ и длина, и угол перистости мышечных волокон уменьшаются, что может указывать на потерю последовательно и параллельно расположенных саркомеров. Функциональным последствием уменьшения длины волокна может быть уменьшение степени укорочения волокна во время сокращения. Потеря последовательно соединенных саркомеров предполагает, что это будет иметь значение для характеристических кривых сила - длина и сила - скорость мышцы. Обнаруженный меньший угол перистости волокна во время сокращения мышцы после иммерсии с одновременным использованием ФЭСТ, очевидно, частично дает компенсацию за потерю силы, которая является постоянным «спутником» гравитационных мышц при разгрузке [2, 3, 31, 36, 38], из-за более эффективной передачи силы к сухожилию, развиваемой волокнами. Уменьшенный начальный угол перистости волокон в покое, вероятно, при сокращении увеличится и уменьшит жесткость сухожилия или мышечно-сухожильного комплекса мышцы, что подтверждается в существенном увеличении ∆Lмышцы ЛИМ (с 0,9 до 3,3 мм после иммерсии) во время сокращения. Это наблюдение подтверждается ранее полученными результатами [40]. Более того, снижение числа последовательно соединенных саркомеров позволяет предположить, что величина сокращения развиваемого волокном будет сниженной. Это предположение хорошо подтверждается данными, полученными ранее в условиях иммобилизации [47].
Толщина МИМ, ЛИМ и КМ тренируемой конечности значительно уменьшилась после иммерсии с ФЭСТ. Этот результат опровергает утверждение, что степень атрофии мышц определяется относительным количеством медленносокращающихся волокон мышцы, т.к. МИМ и ЛИМ имеет относительно более высокий процент быстро сокращающихся волокон [48]. Уменьшение толщины МИМ и ЛИМ в тренируемой конечности вызывает крайнее противоречие, поскольку мышцы во время разгрузки тренировались. Причина этого не ясна и указывает, что для сохранения (или по крайней мере для поддержания) сократительных свойств и архитектуры скелетных мышц во время механической разгрузки требуется специфическая (высокоинтенсивная) тренировка.
С другой стороны, увеличение максимального произвольного суставного момента после иммерсии с применением ФЭСТ позволяет предположить, что ФЭСТ, очевидно, увеличивает афферентный поток [23] в условиях его дефицита при гравитационной разгрузке, вызванном длительной экспозицией, которая может также повысить центральную роль в поддержании и нормализации активности систем управления произвольными движениями (по принципу обратной связи) [1].
Более того, тетаническая электрическая стимуляция, приложенная поверхностно к мышце человека и вызывающая сокращение, деполяризует моторные аксоны, расположенные ниже электродов стимуляции. Одновременная деполяризация сенсорных аксонов также может внести вклад в величину сокращения мышцы через синаптический путь рекрутированием спинальных мононейронов. После входа в спинной мозг сенсорный залп рекрутирует спинальные мононейроны, ведущие к развитию центрального суставного момента. Это рекрутирование совместимо с развитием постоянных внутренних токов в спинальных или межнейронных мотонейронов [9-11]. Постоянные внутренние токи ведут к поддержанию некоторого уровня деполяризации (плато потенциалов), и в связи с этим становится совершенно понятным, что они играют важную роль в регулировании частоты импульсации в нормальных условиях [11, 12, 24]. Максимизирование центрального вклада может быть выгодным для увеличенной силы сокращения мышцы.
В заключение, полученные результаты позволяют сделать вывод, что, во-первых, архитектура разных головок ТМГ значительно разнится, отражая, вероятно, их функциональные роли, во-вторых, различные изменения длины и угла перистости волокон между разными мышцами, вероятно, определяются различиями в способности развивать силу и упругих характеристик сухожилий или мышечно-сухожильного комплекса мышц. Наконец,
в-третьих, ФЭСТ оказывает в целом профилактическое воздействие на стимулируемые мышцы - частично уменьшает глубину и скорость снижения силы сокращения, а также атрофические процессы мышцы, вызванной механической разгрузкой.
Авторы выражают благодарность инженеру Н.Н. Химорода за техническое обеспечение при работе с изокинетическим динамометром.
Особую благодарность авторы выражают всем испытуемым, без их участия которых невозможно было бы получить фактический материал.
СПИСОК ЛИТЕРАТУРЫ
- Бернштейн Н.А. Очерки по физиологии движений и физиологии активности // М.: Медицина, 1966. - 349 с.
- Григорьева Л.С., Козловская И.Б. Влияние 7-суточной иммерсионной гипокинезии на характеристики точностных движений // Космич. биология и авиакосм медицина. - 1985. - №19. - P. 38-42.
- Киренская А.В., Козловская И.Б., Сирота М.Г. Влияние иммерсионной гипокинезии на характеристики ритмической активности двигательных единиц // Физиол. человека. - 1986. - №12. - С. 627-632.
- Козловская И.Б., Григорьева Л.С., Гевлич Г.И. Сравнительный анализ влияний невесомости и ее моделей на скоростно-силовые свойства и тонус скелетных мышц человека // Косм. биология и авиакосм медицина. - 1984. - №6. - С. 22-26.
- Шульжено Е.В., Виль-Вильямс И.Е. Возможности проведения длительной водной иммерсии методом «сухого» погружения // Косм. биология и авиакосм. медицина. - 1976. - №10, - С. 82-84.
- Akima H., Kubo K., Kanehisa H., Suzuki Y., Gunji A., Fukunaga T. Leg-press resistance training during 20 days of 6° head-down-tilt bed rest prevents muscle deconditioning // Eur. J. Appl. Physiol. - 2000 - №82. - P. 30- 38.
- Akima H., Kubo K., Imai M., Kanehisa H., Suzuki Y., Gunji A., Fukunaga T. Inactivity and muscle: effect of resistance training during bed rest on muscle size in the lower limb. // Acta Physiol. Scand. - 2001. - №172. - 269- 278.
- Almekinders L.C. Transcutaneous muscle stimulation for rehabilitation. // Phys. Sportsmedic. - 1984. - №12. - P. 118-124.
- Collins D.F., Burke D., Gandevia S.C. Large involuntary forces consistent with plateau-like behavior of human motoneurons // J. Neurosci. - 2001. - №21. - P. 4059- 4065.
- Collins D.F., Burke D., Gandevia S.C. Sustained contractions produced by plateau-like behaviour in human motoneurones. // J. Physiol. - 2002a. - №538. - P. 289-301.
- Collins D.F., Gorassini M., Bennett D.J., Burke D., Gandevia S.C. Recent evidence for plateau potentials in human motoneurones// In: Sensorimotor Control of Movement and Posture / Eds. Gandevia S.C., Proske U., Stuart D.G. - New York: Plenum. Adv. Exp. Med. Biol. - 2002b. - №508. - P. 227-235.
- Gorassini M., Yang J.F., Siu M., Bennett D.J. Intrinsic activation of human motoneurons: possible contribution to motor unit excitation. // J. Neurophysiol. - 2002. - №87. - P. 1850-1858.
- Cutts А. The range of sarcomere lengths in the muscles of the human lower limb. // J. Anat. - 1988. - №160. - P. 79-88.
- Berg H.E., Larsson L., Tesch P.A. Lower limb skeletal muscle function after 6 weeks of bed rest // J. Appl. Physiol. - 1997. - №82. - P. 182-188.
- Blair E., Erlanger J. A comparison of the characteristics of axons through their individual electrical responses. // Am. J. Physiol. - 1933. - №106. - P. 524-564.
- Bock O. Joint position sense in simulated changed-gravity environments // Aviat. Space Environ. Med. - 1994. - №65. - P. 621-626.
- Burke R.E., Edgerton R.V. Motor unit properties and selective involvement in movement // Exerc. Sport Sci. Rev. - 1975. - №3. - P. 31-81.
- Edgerton V.R., Roy R.R. Response of Skeletal Muscle to Spaceflight // In: Fundamentals of Space Life Sciences / Ed. Churchill S. - Chapt. 7. Malabar. FL. Kreiger. - 1995. - P. 105-120.
- Friedrich J.А., Brand R.А. Muscle fiber architecture in the human lower limb // J. Biomech. - 1990. - №23. - P. 91-95.
- Fukunaga T., Ichinose Y., Ito M., Kawakami Y., Fukunago S, Determination of fascicle length and pennation in a contracting human muscle in vivo // J. Appl. Physiol. - 1997. - №82. - P. 354-358.
- Fukunaga T., Roy R.R., Shellock F.G., Hodgson J.A., Day M.K., Lee P.L., Kwong-Fu H., Edgerton V.R. Physiological cross-sectional area of human leg muscles based on magnetic resonance imaging. // J. Orthop. Res. - 1992. - 10. - P. 926-934.
- Gans C., Bock W.J. The functional significance of muscle architecture - a theoretical analysis // Ergebn. Anat. Entwicklung. - 1965. - №38. - P. 115-142.
- Gazenko O.G., Grigoriev A.I., Kozlovskaya I.B. Mechanisms of acute and chronic effects of microgravity // Physiologist. - 1987. - №30 (Suppl.). - P. S1-S5.
- Heckman C.J., Gorassini M.A., Bennett D.J. Persistent inward currents in motoneuron dendrites: implications for motor output // Muscle Nerve. - 2005. - №31. - P. 135-156.
- Henneman E., Somjen G., Carpenter D.O. Functional significance of cell size in spinal motoneurons // J. Neurophysiol. - 1965. - №28. - P. 560-580.
- Ichinose Y., Kawakami Y., Fukunaga T. In vivo measurement of fascicle arrangement of human vastus lateralis muscle using ultrasound. // XVth Cong. Inter. Soc. Biomech/ / Eds. Häkkinen K., Keskinen K.L., Komi P.V., Mero A. - Gummerus, Jyväskylä, Finland. - 1995. - P. 412-413.
- Ilyina-Kakueva E.I., Portugalov V.V., Krivenkova N.P. Space flight effects on the skeletal muscles of rats // Aviat. Space Environ. Med. - 1976. - №47. - P. 700-703.
- Kawakami Y., Akima H., Kubo K., Muraoka Y., Hasegawa H., Kouzaki M., Imai M., Suzuki Y., Gunji A., Kanehisa H., Fukunaga T. Changes in muscle size, architecture and neural activation after 20 days of bed rest with and without countermeasures // Eur. J. Appl. Physiol. - 2001. - №84. - P. 7-12.
- Kawakami Y., Ichinose Y., Fukunaga T. Architectural and functional features of human triceps surae muscle during contraction // J. Appl. Physiol. - 1998. - №85. - P. 398- 404.
- Kawakami Y., Muraoka Y., Kubo K. Suzuki Y., Fukunaga T. Changes in muscle size and architecture following 20 days of bed rest. // J. Gravit. Physiol - 2000. - №7. - P. 53- 60.
- Koryak Yu. Effects of surface electrostimulation on human skeletal muscle // Proc. 5th Vienna Inter. Workshop of Functional Electrostimulation. - 1995. - P. 297-300.
- Koryak Yu. The effect of 120-days of bed rest with and without countermeasures on the mechanical properties of the triceps surae muscle in young female // Eur. J. Appl. Physiol. - 1998a. V. 78. - P. 128-135.
- Koryak Yu. Electromyographic study of the contractile and electrical properties of the human triceps surae muscle in a simulated microgravity environment // J. Physiol. - 1998b. - №510, - P. 287-295.
- Koryak Yu. Electrically evoked and voluntary properties of the human triceps surae muscle: effects of long-term spaceflights // Acta Physiol. Pharmacol. Bulg. - 2001. - №26. - P. 21-27.
- Koryak Yu. «Dry» immersion induces neural and contractile adaptations in the human triceps surae muscle // Environ. Med. - 2002. - №46. - P. 17-27.
- Koryak Yu. Contractile properties and fatiguability of the human triceps surae muscle after exposure to simulated weightlessness // In: From Basic Motor Control to Functional Rec. III. / Ed. Gantchev N. - University Press. Varna. - 2003. - P. 369- 380.
- Koryak Yu., Kozlovskaya I., Gratchev V. et al. Electromyostimulation (EMS) as a countermeasure of the negative effects of weightlessness to prolonged spaceflights // XII Conference on Space Biol. and Aerospace Med. - 2002. - Р. 486-487.
- Kozlovskaya I.B., Aslanova I.F., Grigorieva L.S., Kreidich Yu.V. Experimental analysis of motor effects of weightlessness // Physiologist. - 1982. - №25 (Suppl.). - P. 49- 52.
- Kozlovskaya I., Dmitrieva I., Grigorieva L. et al. Gravitational mechanisms in the motor sistem. Studies in real and simulated weightlessness // In: Stance and Motion. Facts and Concepts / Eds. Gurfinkel V.S., Ioffe M.Ye., Massion J. - Plenum. - N-Y. - 1988. - P. 37-48.
- Kubo K., Akima H., Kouzaki M., et al. Changes in the elastic properties of tendon structures following 20 days bed rest in humans // Eur. J. Appl. Physiol. - 2000. - №83. - P. 463-468.
- Lieber R.L., Frieden J. Functional and clinical significance of skeletal muscle architecture // Muscle Nerve. - 2000. - №23. - P. 1647-1666.
- Mayr W., Freilinger G., Rafolt R. et al. Functional electrostimulation as a countrermeasure - against muscular atrophy in long-term space flights // BAM. - 2000. - №10. - P. 78
- Narici M.V., Binzoni T., Hiltbrand E. et al. In vivo human gastrocnemius architecture with changing joint angle at rest and during graded isometric contraction // J. Physiol. - 1996. - №496. - P. 287-297.
- Scott S.H., Engstrom C.M., Loeb G.E. Morphometry of human thigh muscles. Determination of fascicle architecture by magnetic resonance imaging // J. Anat. - 1993. - №182. - P. 249-257.
- Solomonow M. External control of the neuromuscular system // IEEE Transactions on Biomed. Engin. - 1984. - №31. - P. 752- 763.
- Sugajima Y., Mitarai G., Koeda M., Moritani T. Characteristic changes of motor unit activity in hip joint flexor muscles during voluntary isometric contraction during water immersion // J. Electromyog. Kinesiol. - 1995. - №6. - P. 83-95.
- Williams P.E., Catanese T., Lucey E.G., Goldspink G. The importance of stretch and contractile activity in the prevention of connective tissue accumulation in muscle // J. Anat. - 1988. - №58. - P. 109-114.
- Yamaguchi G.T., Sawa A.G.U. Moran D.W. Fessler M.J., Winters J.M. А survey of human musculotendon асtuаtor parameters // In: Multiple Muscle Systems. Biomechanics and Movement Organization / Eds: Winters J.S., Woo S.L-Y. - New York. Sрringеr-Vегlаg. - 1990. - P. 717-773.
Библиографическая ссылка
Ю.А. Коряк, М.М. Кузьмина, И.В. Бережинский, В.М. Коваленко ПРОДОЛЖИТЕЛЬНАЯ ЭЛЕКТРОМИОСТИМУЛЯЦИОННАЯ ТРЕНИРОВКА МЫШЦ У ЧЕЛОВЕКА В УСЛОВИЯХ МЕХАНИЧЕСКОЙ РАЗГРУЗКИ ДВИГАТЕЛЬНОГО АППАРАТА И ЕЕ ВЛИЯНИЕ НА АРХИТЕКТУРУ И ФУНКЦИЮ ТРЕХГЛАВОЙ МЫШЦЫ ГОЛЕНИ // Фундаментальные исследования. – 2010. – № 3. – С. 68-87;URL: https://fundamental-research.ru/ru/article/view?id=6658 (дата обращения: 20.04.2024).