



Основной характеристикой абсансной эпилепсии являются разряды пик-волна (spike-wave discharges, SWD). Это спонтанно возникающие генерализованные высокоамплитудные эпизоды электрической активности мозга, сопровождающиеся кратковременным отключением сознания. Согласно современным представлениям абсансная эпилепсия является кортикоталамическим типом эпилепсии. Кортикальный фокус в соматосенсорной коре является ведущим в распространении пик-волновой активности по кортикоталамическим нервным сетям в течение спонтанных абсансных судорог («hot spot theory) [4].
Важным звеном в патогенетических механизмах абсансной эпилепсии является дефицит дофаминергической системы, при этом ведущее значение имеет изменение уровня функционирования дофаминовых рецепторов второго типа (DRD2) [1].
В ранее проведенных генетических исследованиях установлено, что у крыс WAG / Rij генотип локуса TAG 1A (DRD2) может представлять различное сочетание двух аллелей - А1 и А2. В исследованной популяции крыс линии WAG / Rij наиболее часто встречали генотип А1 / А2 (75 % животных), остальные генотипы были более редкими: А2 / А2 - 21 % А1 / А1 - 4 % животных. Целенаправленное скрещивание крыс позволило получить на кафедре морфологии и физиологии человека Башкирского государственного университета две субпопуляции гомозиготных крыс указанной линии [3].
Одним из направлений в анализе электрической активности мозга является использование вейвлет-преобразования [9]. Для анализа частотно-временной структуры разрядов пик-волна у пациентов с абсансной эпилепсией метод был специально модифицирован [2, 6, 7, 8].
Целью нашей работы было исследование с помощью модифицированного вейвлет- преобразования особенностей частотно-временной структуры разрядов пик-волна, зарегистрированных в ЭЭГ двух групп гомозиготных крыс линии WAG / Rij, которые различались представительством аллелей в локусе TAG 1A гена рецептора дофамина второго типа (DRD2).
Для работы были использованы 12 животных с генотипом А1 / А1 по локусу TAG 1А гена DRD2 (условное обозначение данной группы А1А1) и 12 животных с генотипом А2 / А2 по тому же локусу (обозначение - А2А2).
Операции вживления электродов на крысах проводили в стерильных условиях. В качестве наркоза использовали хлоралгидрат в дозе 400 мг / кг. После наркотизации и местной анестезии мягких тканей головы проводили скальпирование и вживление нихромовых электродов эпидурально: во фронтальную кору - поле 6 (AP-+3; LM-3); в теменную кору - поле 2 (AP-0; LM-5); в затылочную кору - поле 17 (AP -6; LM-3) [9]. Индифферентный электрод располагался в кости над мозжечком. После восстановительного периода (7-8 дней) производили регистрацию фоновой ЭЭГ. Запись осуществляли в программе EEGView (А. М. Спиридонов). Использовали электроэнцефалограф Bioskript 2000 (Германия), постоянная времени - 0,3 с, фильтр высокой частоты - 70 Гц, параллельно сигнал поступал в компьютер для дальнейшей обработки.
Анализ частотно-временной структуры разрядов был проведен с помощью вейвлет-анализа. Суть преобразования состоит в разложении по базису одномерного сигнала, сконструированного из обладающих определенными свойствами функций, посредством масштабных изменений и переносов.
Непрерывное вейвлет-преобразование W функции f задается формулой Wf (a, b) =1 / √ ∫ f (x) ψ { (x-b) / a} dx, где a, b - коэффициенты масштабирования и сдвига соответственно [2]. Коэффициент b отвечает за сдвиг по времени, а коэффициент а - за частотную характеристику. Для частотно-временного анализа часто используют вейвлет Морле, поскольку он имеют наилучшую одновременно частотную и временную локализацию. Вейвлет Морле определяется следующим выражением: ψ (x) =C1 exp (-x² / 2) cos (C2 x), где C1, C2 - константы. Спектрограммы Морле дают представление об изменении амплитуды в течение разряда, однако точная количественная оценка изменений частоты с помощью обычного вейвлет-преобразования Морле является сложной.
Bosnyakova D, Obukhov Yu., (2005), Габова и др. (2008) модифицировали вейвлет Морле таким образом, чтобы разложение исходного сигнала по модифицированному базису позволяло выделить динамику частоты разряда: ψ (x) =xe-х2 / 2cos (5х). Преобразование сигнала по такому модифицированному базису будет иметь нули в точках, в которых трехмерный «хребет», полученный с помощью обычного преобразования Морле, имеет максимум [2, 6, 7, 8].
Кроме исследования частотно-временной структуры разрядов, был рассчитан пик-волновой индекс. Пик-волновой индекс (SWDInd) - это процент времени, занятый разрядами. Для расчета индекса находили сумму длительности всех пик-волновых разрядов на протяжении всего файла, за 100 % принимали продолжительность файла.
Особенности частотно-временной структуры пик-волновых разрядов крыс группы А1А1. Средняя продолжительность разрядов, зарегистрированных в ЭЭГ крыс группы А1А1, составила 5,6±0,34 с., среднее количество пик-волновых разрядов в проанализированных файлах крыс анализируемой группы равно 8,6±0,66. Еще один важный количественный показатель пик-волновых разрядов, отражающий, какую долю занимают пик-волновые разряды в общей длительности записи, - это пик-волновой индекс. Этот показатель составляет 9,5±0,82 %.
Применяя модифицированное вейвлет преобразование и вейвлет преобразование Морле, мы построили более тысячи преобразований входного сигнала. На рис. 1 представлены результаты вейвлет-преобразования одного из разрядов крыс группы А1А1, зарегистрированного в теменной области коры. Данный пик-волновой разряд имеет продолжительность 4,5 секунды (рис. 1а). Частота комплексов «пик-волна» в начальный момент времени составляет 14,8 Гц, (рис. 1в), в течение первой секунды частота разряда снижается до отметки 8 Гц, а затем на фоне общего небольшого снижения отмечены периодические флуктуации частоты в диапазоне десятых долей герца. Такие флуктуации в течение разряда встречаются несколько раз, и их период составляет около трех секунд (рис. 1в). Средняя начальная частота комплексов «пик-волна» у крыс группы А1А1 равна 14,1±0,10 Гц. Анализируя вейвлет-преобразование Морле (рис. 1б), мы отметили, что коэффициент вейвлета - W - (пропорционален амплитуде) колеблется в диапазоне от 1000 до 2000 мкВ.
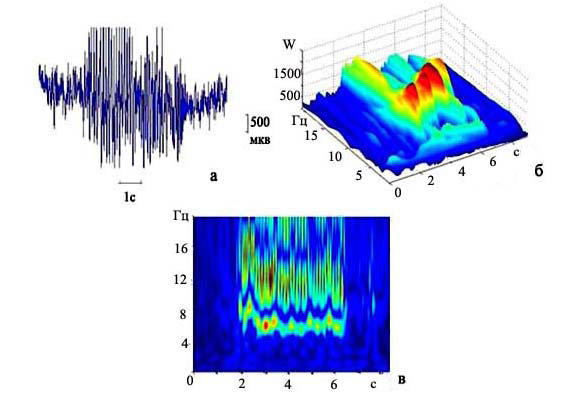
Рис. 1. Частотно-временная структура пик-волнового разряда крыс группы А1А1; а) запись разряда пик-волна, калибровка 1с., 500 мкВ; б) трехмерный график преобразования Морле данного разряда. По осям на трехмерном графике - время (с.), частота (Гц), коэффициент вейвлета (W); в) анализ с помощью модифицированного вейвлета: по горизонтали - время, по вертикали - частота
Особенности частотно-временной структуры пик-волновых разрядов крыс группы А2А2. Средняя продолжительность разрядов у крыс данной группы равна 3,72±0,14 с., часто продолжительность разрядов не превышала 2-3 с. Количество пик-волновых разрядов крыс группы А2А2 составила 4,21±0,41. На основе этих параметров нами был посчитан третий количественный показатель пик-волновых разрядов - пик-волновой индекс, и у крыс данной группы он составляет 2,71±0,25 %.
Для анализа частотно-временной структуры разрядов были использованы модифицированное вейвлет преобразование и вейвлет-преобразование Морле. На рис. 2 представлен короткий разряд, характерный для анализируемой группы. Разряд длится около двух секунд. Он начинается с высокой частоты - 14-16 Гц (рис. 2в). В течение всего короткого разряда происходило стремительное снижение его частоты. Скорость уменьшения частоты у коротких разрядов обычно была 1 Гц / с или более. Иногда на этом фоне наблюдали небольшое временное повышение частоты (рис. 2б - на отметке 1,5 с можно наблюдать незначительный подъем частоты). Средняя начальная частота комплексов «пик-волна» у крыс группы А2А2 составила 12,8±0,13 Гц. Коэффициент вейвлета (W) у крыс данной группы колеблется в диапазоне 1000-1500 (рис. 2в).
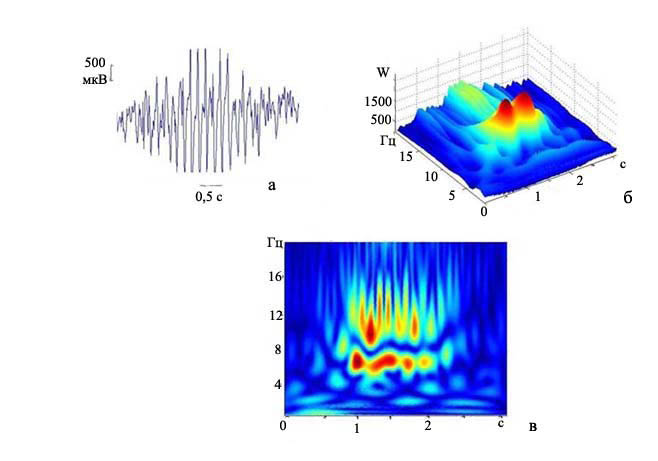
Рис. 2. Частотно-временная структура коротких разрядов, характерных для группы А2А2: а) запись разряда пик-волна, калибровка 0,5с., 500 мкВ; б) трехмерный график преобразования Морле данного разряда. По осям на трехмерном графике - время (с.), частота (Гц), коэффициент вейвлета (W); в) анализ с помощью модифицированного вейвлета: по горизонтали - время, по вертикали - частота.
Проанализировав полученные вейвлет-преобазования, мы обнаружили, что у крыс группы А2А2 имеются разряды, которые не подходят по описанию тем разрядам, которые были охарактеризованы выше. Для них свойственна менее выраженная симметричность по сравнению с вышеописанными разрядами, они - асимметричны. В большинстве случаев не наблюдали четкого опережения разряда во фронтальной коре. Максимальная частота комплексов «пик-волна» могла приходиться как на середину, так и на его конец. Эти разряды были обозначены как «атипичные». На рис. 3 представлен вейвлет-анализ одного из «атипичных» разрядов, зарегистрированных нами у крыс группы А2А2. Максимальная частота комплексов «пик-волна» приходится на середину разряда (рис. 3в), коэффициент вейвлета (W) в начальный момент разряда составляет 1500, затем происходит значительное колебание его значения, которое выражается образованием двух «хребтов» на вейвлет преобразовании Морле (рис. 3б, показано стрелочками). Для крыс группы А2А2 мы отметили чередование широкогенерализованных по коре разрядов с разрядами с «атипичной» структурой.
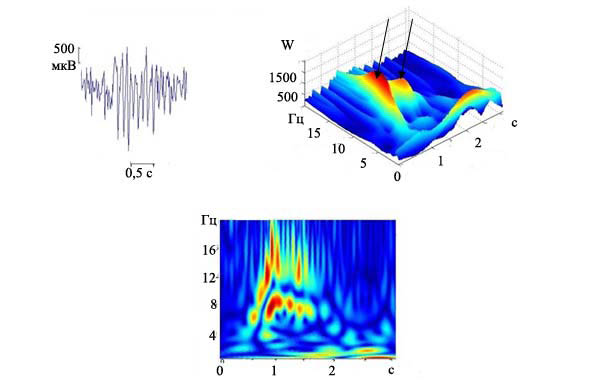
Рис. 3. Частотно-временная динамика атипичного разряда, зарегистрированного в ЭЭГ группы А2А2: а) запись разряда пик-волна, калибровка 0,5с., 500 мкВ; б) трехмерный график преобразования Морле данного разряда. По осям на трехмерном графике - время (с.), частота (Гц), коэффициент вейвлета (W); в) анализ с помощью модифицированного вейвлета: по горизонтали - время, по вертикали - частота
В настоящей работе были исследованы особенности структуры одного из видов высокосинхронизированной патологической активности коры головного мозга - разрядов «пик-волна» неконвульсивной абсансной эпилепсии. Для крыс группы А1А1 были получены следующие результаты: в самом начале разряда «пик-волна» частота была наиболее высокой в лобных отделах коры, и этот начальный период разряда всегда был очень коротким. Эти данные согласуются с ранее проведенными исследованиями на крысах линии WAG / Rij [2, 6, 7]. Картина изменений частоты коротких разрядов (менее 4 с.), свойственных группе А2А2, несколько отличается от описанной у крыс группы А1А1. У таких разрядов снижение частоты происходит стремительно. На ЭЭГ группы А2А2 одновременно присутствовали как «типичные», так и «атипичные» разряды. Такие «атипичные разряды» были описаны у пациентов с неконвульсивной эпилепсией, была выявлена их структурная неоднородность [2].
Выводы
1. Крысы группы А1А1 имеют достоверно большую продолжительность SWD в ЭЭГ соматосенсорной коры по сравнению с крысами группы А2А2; p<0,001. Количество SWD в ЭЭГ крыс группы А1А1 значимо больше по сравнению с группой А2А2 (p<0,001).
2. Пик-волновой индекс (время, занятое разрядами) у крыс группы А2А2 достоверно ниже по сравнению с тождественным показателем у группы А1А1; p<0,001.
3. По частотно-временной структуре крысы группы А1А1 и А2А2 неоднородны: для группы А1А1 характерны типичные разряды, для группы А2А2 - одновременное присутствие типичных и атипичных разрядов на ЭЭГ.
СПИСОК ЛИТЕРАТУРЫ
- Бирюкова Л. М., Мидзяновская И. С., Ленсу С., Туомисто Л., Ван Луийтелаар Ж., Базян А. С.// Нейрохимия. - 2006; 23: 3:234-239.
- Габова А. В., Гнездицкий В. В., Боснякова Д. Ю., Жарикова А. В., Самотаева И. С., Обухов Ю. В., Кузнецова Г. Д. Технологии живых систем, 2008; 5: 72-81.
- Калимуллина Л. Б., Ахмадеев А. В., Бикбаев А. Ф., Баязитова Л. И. // Материалы V съезда Российского общества медицинских генетиков. - Уфа // Медицинская генетика. - 2005. - № 4. - С. 150.
- Меерен Х. К. М., Е. Л. Дж. М. Ван Луителлаар, Ф. Х. Лопес да Сильва, Р. К. Бердиев, Н. Е. Чепурнова, С. А. Чепурнов, А. М. Л. Кунен. // Успехи физиологических наук. - 2004. - Т. 35, № 1. - С. 3-19
- Мидзяновская И. С., Кузнецова Г. Д., Туомисто Л..и др. // Нейрохимия. - 2004. - Т. 21, № 4. - С. 264-270.
- Acar T., Aykut-Bingol C., Bingol H., Bro R., Yener B. // Bioinformatics 2007; 23: 110-118.
- Bosnyakova D., Gabova A., Zharikova A., Gnezditski V., Kuznetsova G., van Luijtelaar G. // Clin. Neurophysiol. 2007; 118: 1736-1743.
- Malat S. G. Wavelet tour for signal processing. - San Diego: Academic Press. 1999.
- Paxinos G., Watson C. The rat brain in stereotaxic coordinates. - Sydney: Academic, 1998.
Библиографическая ссылка
А.М. Мусина, Л.Б. Калимуллина ЧАСТОТНО-ВРЕМЕННАЯ ХАРАКТЕРИСТИКА ПИК-ВОЛНОВЫХ РАЗРЯДОВ В ЭЛЕКТРОЭНЦЕФАЛОГРАММЕ КРЫС С РАЗЛИЧИЯМИ В АЛЛЕЛЬНОЙ СТРУКТУРЕ ГЕНА РЕЦЕПТОРА ДОФАМИНА ВТОРОГО ТИПА (DRD2) // Фундаментальные исследования. – 2010. – № 6. – С. 7-13;URL: https://fundamental-research.ru/ru/article/view?id=8949 (дата обращения: 23.04.2024).