



Основой изучения и сохранения глобального биоразнообразия является инвентаризация видового состава региональных биот с целью выяснения механизмов и условий их устойчивого функционирования. Несмотря на значительные усилия по инвентаризации биоразнообразия регионов России, во многих регионах уровень изученности видового состава и структурных особенностей биоты недостаточен. В особенности это касается групп живых организмов, которые по тем или иным причинам остаются аутсайдерами с точки зрения объектов исследований. К таким организмам, в частности, относятся грибы, являющиеся неотъемлемой частью системы редуцентов.
Роль редуцентов и в особенности деструкторов в экосистемах исключительна. Деструкторы и детритофаги являются главными потребителями растений на суше; они обеспечивают переработку мортмассы и возвращение биогенных элементов в круговорот, в чем крайне «заинтересованы» экосистемы [13]. Кроме того, они являются вторичными продуцентами, обеспечивают перераспределение органического вещества и косвенно влияют на изменение структуры фитоценозов [1]. Основную часть системы редуцентов составляют бактерии, грибы и беспозвоночные. Часто эти организмы совместно осуществляют механическое и химическое разрушение детрита, но в определенных условиях, например, в лесных экосистемах на первое место по объемам осваиваемого детрита выходят базидиальные грибы, которые способны самостоятельно, практически без участия других организмов, осуществлять полную деструкцию растительной органики.
Изучению особенностей биологии этой группы грибов уделяется значительное внимание в современной микологической науке, однако многие регионы остаются «белыми пятнами» с точки зрения выявления видового состава ксилотрофных базидиомицетов, их экологии, закономерностей их распространения и особенностей функционирования их сообществ.
В отношении грибов-макромицетов являются актуальными все проблемы, связанные с выявлением их разнообразия и динамики, характерные в целом для растительного царства, в частности, проблемы полноты выявления видового состава, применения метода локальных флор и т.п. [11, 12 и др.]. Необходимость применения особых подходов к оценке разнообразия споровых растений отмечал Б.А. Юрцев [12], указывавший, что они отличаются от прочей флоры намного более обширными ареалами, но сильнее дифференцированы по микронишам.
Материалы и методы исследования
В рамках работ по исследованию структурных особенностей биоты грибов-макромицетов Южного Приуралья в пределах Оренбургской области [2], с 1994 по 2012 гг. проводилось ежегодное изучение видового состава дереворазрушающих базидиомицетов, обитающих в лесах предгорий Южного Урала (Тюльганский район Оренбургской области). Здесь представлены широколиственные леса с доминированием Quercus robur L., Ulmus laevis Pall., Acer platanoides L., Tilia cordata Mill.; по распадкам и в нижней части пологих склонов широко распространены березняки и осинники; в поймах рек и ручьев – ольшаники с доминированием Alnus glutinosa (L.) Gaertn., A.incana (L.) Moench, ивняки и тополевники. Относительно невысокий уровень антропогенной нагрузки на леса района, обусловленный спецификой размещения населения и отсутствием подъездных путей, определяет повышенный интерес к изучению всех компонентов этих лесных экосистем и, в частности, грибов-макромицетов. Большая часть работ проводилась в лесах в окрестностях с. Ташла в радиусе 10 км от села; чаще всего исследования проводились в конце июня – начале июля.
Результаты исследования и их обсуждение
В итоге проведенных исследований было учтено 2722 плодовых тела, идентифицированных как 189 видов, относящихся к 86 родам, 30 семействам и 19 порядкам базидиальных дереворазрушающих грибов. В ходе исследований мы отметили существенное варьирование видового разнообразия локальной микобиоты, а также явную неравномерность встречаемости отдельных видов по годам.
Можно было бы предположить, что проведение методически одинаковых многолетних исследований видового состава биоты должно обеспечить выявление полного состава видов, однако анализ (рис. 1) показывает устойчивую тенденцию к повышению общего богатства микобиоты.
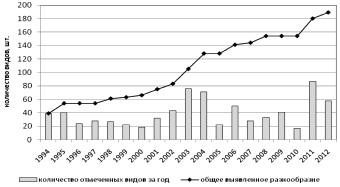
Рис. 1. Изменение видового разнообразия локальной микобиоты по годам
Отмеченное нарастание общего выявленного разнообразия происходит на фоне периодических колебаний числа выявленных видов. При этом меняется не только видовой состав грибов в изучаемых лесных экосистемах, но столь же регулярно варьируется и количество отмеченных плодовых тел (коэффициент корреляции Спирмена – 0,73) (рис. 2).
Таким образом, наблюдается периодическое варьирование как видового разнообразия, так и относительной численности видов с периодом 10 лет. Полагая, что эта периодичность должна быть обусловлена периодичностью влияния факторов среды, мы провели корреляционный анализ изменчивости показателей микобиоты и климатических показателей района исследований, которые могли бы оказать существенное влияние на развитие грибов и формирование плодовых тел, а именно: температуры мая, июня, июля, августа; средняя температура теплых месяцев, среднемесячная температура за год; температура максимально теплого месяца; относительная влажность за теплые месяцы; относительная влажность за год; минимальная, максимальная, средняя годовая температура (данные архива http://thermograph.ru).
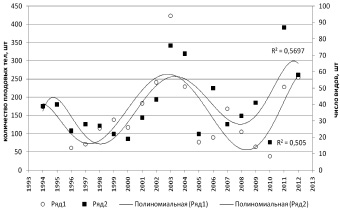
Рис. 2. Варьирование количества плодовых тел и видового разнообразия по годам наблюдений (аппроксимация полиномиальной функцией). Ряд 1 – количество плодовых тел; Ряд 2 – число видов
Исходя из общепринятых воззрений на экологические потребности грибов, можно было бы предположить, что ключевым фактором, определяющим их разнообразие и встречаемость, является относительная влажность и количество осадков. Однако проведенные ранее в регионе исследования показали, что у разных видов грибов ключевые факторы отличаются [5]. В рамках проводимого исследования корреляция между количеством осадков и характеристиками микобиоты также была не значима (коэффициент корреляции менее 0,5). Это выглядит вполне логичным, если учесть сложность рельефа района исследований, разную крутизну и экспозицию склонов, различия конкретных лесорастительных условий и характеристик древостоев и т.п., что определяет широкое варьирование показателей увлажнения по территории.
Наиболее достоверная отрицательная корреляция (–0,65) была выявлена между видовым разнообразием микобиоты и температурой максимально теплого месяца. Можно предположить, что высокие температуры препятствуют функционированию в субстрате мицелиев многих видов и они не имеют возможности формировать плодовые тела.
Между комплексами видов разных лет отмечено достаточно высокое сходство (средняя преемственность видового состава составляет 45,9 %). Соответственно, некоторые виды должны постоянно отмечаться в микобиоте, а появление других должно подчиняться закономерностям изменения условий среды или появляться в годы с нетипичными условиями.
Исходя из встречаемости в ходе многолетнего мониторинга, мы выделили несколько групп видов. Первая из них – виды, являющиеся постоянными компонентами микобиоты; к этой группе можно отнести 13,8 % видов. Это виды с высоким потенциалом выявления [3], с тримитической или димитической гифальной системой, формирующие крупные и/или многолетние базидиомы, в частности, Bjerkandera adusta, Cerrena unicolor, Fomes fomentarius, Fomitopsis pinicola Ganoderma lipsiense, Phellinus igniarius, Polyporus squamosus, виды рода Trametes и др. Постоянное присутствие в микоценозах некоторых других видов, вероятно, определяется постоянным наличием предпочитаемых ими субстратов (Daedalea quercina, Fomitoporia robusta, Hymenochaete tabacina, Phellinus alni, Phellinus tremulae, Piptoporus betulinus и др.).
12,7 % видов отмечались в микобиоте периодически; появление их базидиом или значительное возрастание численности, по-видимому, следует связывать с периодичностью климатических показателей. К ним, в частности, относятся Daedaleopsis septentrionalis, Laetiporus sulphureus, Lenzites betulina, Polyporus arcularius, Skeletocutis nivea, Tyromyces chioneus и др.
Значительный интерес представляет малочисленная группа видов, которые были отмечены только в периоды «пиков» видового разнообразия микобиоты. К ним, в частности, относятся некоторые агарикоидные дереворазрушающие грибы (виды рода Lentinus), а также Ceriporia viridians, Inocutis rheades, Inonotus radiatus, Phlebia tremellosa, Spongipellis spumeus и ряд других видов, которые, судя по всему, требуют более оптимальных условий увлажнения для развития базидиом.
И, наконец, 34,4 % видов могут быть отнесены к малочисленным, т.е. за все время исследований они были представлены единичными находками. Вероятно, эти или случайные виды, не типичные для локальной микобиоты (виды родов Antrodiella, Ceriporiopsis, Hyphoderma, Hyphodontia, Pluteus, Postia, Steccherinum) или редкие, требующие особого внимания в целях определения их природоохранного статуса (Hericium coralloides, Ischnoderma resinosum, Pycnoporellus fulgens, Sarcodontia crocea, Tyromyces fumidiceps, T. kmetii) [9].
Обсуждая возможные причины «волн» видового разнообразия в рассматриваемой микобиоте, необходимо учесть, что помимо климатических факторов, влияние которых на видовую структуру было рассмотрено выше, на разнообразие влияют и широкий спектр иных – эндогенных и экзогенных факторов. В частности, различия видового состава могут быть связаны с сукцессионными преобразованиями лесных экосистем и их микоценозов. Влияние разных типов сукцессий на видовой состав и структуру сообществ грибов не однотипно и зависит как от причин сукцессий, так и от структуры конкретных древостоев. Наши исследования охватывали значительную территорию, в пределах которой происходили дигрессии и демутации сообществ разных типов (эксплуатациогенные, пирогенные, зоогенные и др.) [4, 7]. Тенденции изменения видового состава в рамках этих сукцессий отличаются [8, 10], что в итоге создает сложную картину динамики микобиоты, из которой трудно четко вычленить отдельные процессы.
Воспринимая картину динамики локальной микобиоты в целом, можно сделать практический вывод, направленный на оптимизацию системы сохранения биоразнообразия лесных экосистем региона и микобиоты в частности. Для получения объективной картины видового богатства и выявления местообитаний редких и малочисленных видов в лесных массивах, перспективных для создания особо охраняемых территорий, необходимо проведение в них целенаправленных микологических исследований как минимум в течение 5 лет, с тем, чтобы выявить достоверный уровень видового разнообразия. В этом случае в поле зрения специалистов попадет большая часть видов и создаваемая модель ООПТ станет более адекватной [6].
Исследования выполнены при финансовой поддержке гранта губернатора и правительства Оренбургской области «Комплексная экологическая оценка состояния биоты искусственных лесных насаждений Оренбургского Предуралья» (2013 г.)
Рецензенты:
Мирошников С.А., д.б.н., профессор, директор Всероссийского НИИ мясного скотоводства, г. Оренбург;
Паршина Т.Ю., д.б.н., доцент, профессор кафедры зоологии, экологии и анатомии, ФГБОУ ВПО «Оренбургский государственный педагогический университет», г. Оренбург.
Работа поступила в редакцию 22.07.2013.
Библиографическая ссылка
Сафонов М.А., Сафонова Т.И., Каменева И.Н. МНОГОЛЕТНЯЯ ДИНАМИКА ВИДОВОЙ СТРУКТУРЫ ЛОКАЛЬНОЙ МИКОБИОТЫ В ЛЕСАХ ПРЕДГОРИЙ ЮЖНОГО УРАЛА // Фундаментальные исследования. – 2013. – № 10-3. – С. 575-579;URL: https://fundamental-research.ru/ru/article/view?id=32322 (дата обращения: 20.04.2024).