



Одной из характерных черт степной зоны Евразии является ее малая лесистость, определяющаяся аридными и субаридными условиями климата, особенностями почвенного покрова и рядом других причин, связанных с самим генезисом данного типа растительности. Территория Оренбургской области, расположенной на юго-востоке европейской части России, представляет собой большой массив степей и сельскохозяйственных земель на их месте, которые к северу сменяются лесостепными ландшафтами.
Травянистые сообщества занимают основные пространства региона, оставляя лесам понижения рельефа, берега рек. Также лесные сообщества встречаются на склонах возвышенности Общий Сырт, а также представлены достаточно многочисленными насаждениями антропогенного происхождения. Низкая лесистость региона определила тип его хозяйства и при этом повышенный интерес к лесным насаждениям, вычленяющимся из общей структуры ландшафтов и биоты территории. Лесная растительность в пределах степной зоны создает многочисленные экотонные зоны, в которых наблюдается пик биоразнообразия вследствие смешения в них элементов биоты с разными экологическими преференциями; лесные экосистемы являются рефугиумами для ряда видов, в целом не типичных для степей.
Вместе с давлением неблагоприятных факторов среды, леса степных районов испытывают и сильную антропогенную нагрузку из-за их активного и часто нерегулируемого использования в качестве объектов рекреации, побочного и основного лесопользования, техногенных и зоогенных нарушений прилегающих экосистем и т.п. В связи с этим встает вопрос о необходимости анализа современного состояния экосистем этих лесов с целью определения основных путей их развития; разработки и апробации мер, объективно способствующих их сохранению. В этом плане, одной из основных задач является выявление устойчивых лесов степных районов на базе формирования полноценного кадастра биоты, включающего все основные группы живых организмов.
Одной из групп, сравнительно мало изученных в Оренбургском Предуралье, являются грибы-макромицеты и, в частности, древоразрушающие базидиальные грибы, составляющие неотъемлемую часть системы редуцентов лесных экосистем, в том числе и в пределах степной зоны Южного Урала.
Материалы и методы исследований
Объектом исследования были древоразрушающие (ксилотрофные) базидиальные грибы, обитающие на древесине разной видовой принадлежности и находящейся на разной стадии разложения. Для изучения биоты древоразрушающих грибов в 1993–2014 гг. был обследован ряд насаждений разного видового состава в степных районах центральной части Оренбургской области (рис. 1). В общей сложности были обследованы искусственные насаждения на площади более 120 га. Площадь отдельных обследованных площадок колебалась от 0,25 до 2 га, исходя из площади, реально занимаемой тем или иным насаждением.
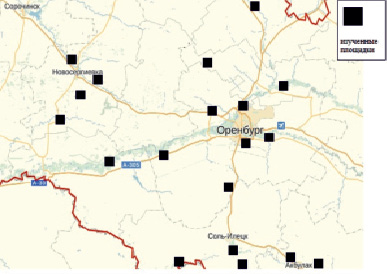
Рис. 1. Карта изученных лесных массивов в степной зоне центральной части Оренбургской области
Сбор образцов производился методом маршрутного учета. На маршруте производилось описание биотопов, растительности и субстратов, на которых обитали грибы. В общей сложности было собрано и определено более 1200 образцов плодовых тел. Идентификация собранных образцов была произведена автором с использованием русскоязычной и зарубежной определительной литературы. При анализе использовались определенные и не идентифицированные ранее материалы, хранящиеся в коллекции ФГБОУ ВПО «Оренбургский государственный педагогический университет».
При описании грибов была использована система высших базидиальных грибов, опубликованная в книге «Nordic Macromycetes» [8, 9].
Результаты исследования и их обсуждение
В результате многолетних исследований биоты древоразрушающих грибов степных районов Оренбургского Предуралья было выявлено 126 видов древоразрушающих грибов, относящихся к 68 родам и 32 семействам отдела Basidiomycota. Большая их часть относилась к афиллофороидным и кортициоидным грибам; меньше представлены в изученной биоте агарикоидные грибы, что связано со спецификой исследований по изучению биоты ксилотрофных грибов Южного Приуралья.
Ведущими семействами биоты древоразрушающих грибов являются Coriolaceae, Fomitopsidaceae, Phellinaceae, Polyporaceae, что характерно в целом для микобиоты ксилотрофных грибов региона [5]. Сравнительный анализ биоразнообразия грибов в разных частях района исследования показал, выявленные виды распределены по изученной территории неравномерно (таблица).
Распространение видов древоразрушающих грибов в районах исследований*
|
Виды грибов |
Районы исследования |
||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
|
Abortiporus biennis (Bull.: Fr.) Singer |
+ |
+ |
|||||
|
Amphinema byssoides (Fr.) J.Erikss. |
+ |
||||||
|
Antrodia serialis (Fr.) Donk |
+ |
||||||
|
Armillaria mellea (Vahl.:Fr.) Kumm. |
+ |
||||||
|
Auricularia mesenterica (Gmel.: Fr.) Pers. |
+ |
||||||
|
Bierkandera adusta (Willd.: Fr.) P.Karst. |
+ |
+ |
|||||
|
Calocera viscosa (Pers.:Fr.) Fr. |
+ |
+ |
|||||
|
Ceraceomyces eludens K.H.Larss. |
+ |
||||||
|
Ceriporia reticulate (Hoffm.:Fr.) Domański |
+ |
||||||
|
Cerrena unicolor (Bull.: Fr.) Murrill |
+ |
+ |
+ |
+ |
+ |
||
|
Chondrostereum purpureum (Pers.: Fr.) Pouzar |
+ |
+ |
+ |
+ |
|||
|
Clavicoruna pixidata (Pers.: Fr.) Doty |
+ |
||||||
|
Coniophora arida (Fr.) P.Karst. |
+ |
||||||
|
Coniophora olivaceae (Pers.:Fr.) P.Karst. |
+ |
||||||
|
Crepidotus luteolus (Lambotte) Sacc. |
+ |
||||||
|
Cylindrobasidium evolvens (Fr.: Fr.) Jülich |
+ |
||||||
|
Cylindrobasidium laeve (Pers.:Fr.) Chamuris |
+ |
||||||
|
Dacryobolus sudans (Alb. & Schwein.: Fr.) Fr. |
+ |
||||||
|
Daedalea quercina (L.: Fr.) Pers. |
+ |
+ |
+ |
+ |
+ |
||
|
Daedaleopsis confragosa (Bolton: Fr.) Schroet. |
+ |
+ |
+ |
||||
|
Daedaleopsis tricolor (Pers.) Bondartsev & Singer |
+ |
||||||
|
Datronia mollis (Sommerf.: Fr.) Donk |
+ |
||||||
|
Datronia stereoides (Fr.) Ryvarden |
+ |
+ |
+ |
||||
|
Diplomitoporus flavescens (Bres.) Ryvarden |
+ |
+ |
|||||
|
Exidia glandulosa (Bull.: Fr.) Fr. |
+ |
+ |
+ |
+ |
|||
|
Fistulina hepatica (Schaeff.: Fr.) Fr. |
+ |
+ |
+ |
||||
|
Flammulina velutipes (Curt.: Fr.) Singer |
+ |
||||||
|
Fomes fomentarius (L.: Fr.) Fr. |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Fomitoporia punctata (P.Karst.) Pilat |
+ |
+ |
|||||
|
Fomitoporia robusta (P.Karst.) Fiasson & Niemelä |
+ |
+ |
|||||
|
Fomitopsis pinicola (Sowerby: Fr.) P.Karst. |
+ |
+ |
+ |
||||
|
Ganoderma lipsiense (Batsch.) G.F.Atk. |
+ |
+ |
+ |
+ |
+ |
+ |
|
|
Gloeophyllum sepiarium (Wulfen: Fr.) P.Karst. |
+ |
||||||
|
Gloeoporus dichrous (Fr.) Bres. |
+ |
+ |
|||||
|
Gloeoporus taxicola (Pers.: Fr.) Gilb. & Ryvarden |
+ |
+ |
+ |
||||
|
Hapalopilus rutilans (Pers.: Fr.) P.Karst. |
+ |
+ |
+ |
||||
|
Hymenochaete tabacina (Fr.) Lev. |
+ |
+ |
|||||
|
Hyphoderma guttuliferum (P.Karst.) Donk |
+ |
||||||
|
Hyphoderma praetermissum (P.Karst.) J.Erikss. & A.Strid |
+ |
||||||
|
Hyphoderma puberum (Fr.) Wallr. |
+ |
||||||
|
Hyphodontia breviseta (Karst.) Eriksson |
+ |
||||||
|
Hyphodontia crustosa (Pers.: Fr.) J.Erikss. |
+ |
||||||
|
Hyphodontia nespori (Bres.) J.Erikss. & Hjortstam |
+ |
||||||
|
Hyphodontia sambuca (Pers.: Fr.) J.Erikss. |
+ |
||||||
|
Hypholoma fasciculare (Huds.: Fr.) Kumm. |
+ |
||||||
|
Hypholoma Candoleanum (Fr.) Quel. |
+ |
||||||
|
Hypholoma sublateretium(Fr.) Quel. |
+ |
+ |
|||||
|
Hypochnicium vellereum (Eliss. & Cragin) Parmasto |
+ |
||||||
|
Inocutis dryophila (Berk.) Fiasson & Niemelä |
+ |
+ |
+ |
||||
|
Inocutis rheades (Pers.) Fiasson & Niemelä |
+ |
+ |
+ |
+ |
+ |
+ |
|
|
Inonotus obliquus (Pers.: Fr.) Pilat |
+ |
||||||
|
Inonotus radiatus (Sowerby: Fr.) P.Karst. |
+ |
||||||
|
Irpex lacteus (Fr.) Fr. |
+ |
+ |
+ |
+ |
|||
|
Laetiporus sulphureus (Bull.: Fr.) Murrill |
+ |
+ |
+ |
+ |
|||
|
Lentinus adhaerens (Alb. & Schw.: Fr.) Fr. |
+ |
+ |
|||||
|
Lentinus cyathiformis (Schaeff.) Bres. |
+ |
+ |
|||||
|
Lentinus tigrinus (Bull.: Fr.) Fr. |
+ |
+ |
|||||
|
Lenzites warnieri Dur. & Mont |
+ |
+ |
+ |
+ |
|||
|
Leucogyrophana mollusca (Fr.) Pouzar |
+ |
||||||
|
Oxyporus corticola (Fr.) Ryvarden |
+ |
+ |
+ |
+ |
+ |
||
|
Oxyporus obducens (Pers.) Donk |
+ |
+ |
+ |
||||
|
Panellus stipticus (Bull.: Fr.) P.Karst. |
+ |
+ |
|||||
|
Peniophora limitata (Chaill.: Fr.) Cooke |
+ |
||||||
|
Peniophora pini (Schleich.: Fr.) Boidin |
+ |
||||||
|
Peniophorella tsugae (Burt) K.H.Larss. |
+ |
||||||
|
Phanerochaete calotricha (P.Karst.) J.Erikss. & Ryvarden |
+ |
||||||
|
Phanerochaete laevis (Pers.: Fr.) J.Erikss. & Ryvarden |
+ |
||||||
|
Phanerochaete sanguinea (Fr.) Pouzar |
+ |
||||||
|
Phellinus alni (Bondartsev) Parmasto |
+ |
||||||
|
Phellinus igniarius Niemelä s.lato |
+ |
+ |
+ |
+ |
+ |
+ |
|
|
Phellinus linteus (Berk. et Curt.) Teng |
+ |
+ |
+ |
+ |
+ |
||
|
Phellinus rhamni (M.Bond.) Jahn. |
+ |
||||||
|
Phellinus tremulae (Bondartsev) Bondartsev & Borisov |
+ |
+ |
|||||
|
Phellinus tuberculosus (Baumg.) Niemelä |
+ |
||||||
|
Phlebia albida H. Post |
+ |
||||||
|
Phlebia cornea (Bourdot & Galzin) Parmasto |
+ |
||||||
|
Phlebia rufa (Pers.: Fr.) M.P.Christ. |
+ |
||||||
|
Phlebia tremellosa (Schrad.: Fr.) Burds. & Nakasone |
+ |
||||||
|
Phlebia tristis (Litsch. & S.Lundell) Parmasto |
+ |
||||||
|
Pholiota aurivella (Batsch: Fr.) Kumm. |
+ |
||||||
|
Pholiota destruens (Bond.) Gill. |
+ |
+ |
|||||
|
Pholiota squarrosoides (Peck.) Sacc. |
+ |
||||||
|
Piptoporus betulinus (Bull.: Fr.) P.Karst. |
+ |
+ |
+ |
||||
|
Pleurotus calyptratus (Lindbl.) Sacc. |
+ |
+ |
+ |
+ |
+ |
||
|
Pleurotus dryinus (Pers.: Fr.) Kumm. |
+ |
+ |
|||||
|
Pleurotus ostreatus (Jacq.: Fr.) Kumm. |
+ |
+ |
+ |
||||
|
Pleurotus pulmonarius (Fr.) Quel. |
+ |
+ |
|||||
|
Pluteus atricapillus (Batsch.) Fayod |
+ |
+ |
|||||
|
Pluteus tomentosulus Peck. |
+ |
||||||
|
Polyporus arcularius Batsch.: Fr. |
+ |
+ |
+ |
||||
|
Polyporus squamosus Huds.: Fr. |
+ |
+ |
+ |
+ |
|||
|
Polyporus varius (Pers.) Fr. |
+ |
||||||
|
Porodaedalea conchata (Pers.: Fr.) Fiasson & Niemelä |
+ |
||||||
|
Postia caesia (Shrad.: Fr.) P.Karst |
+ |
||||||
|
Postia fragilis (Fr.) Jülich |
+ |
||||||
|
Postia hibernica (Berk.& Broome) Jülich |
+ |
||||||
|
Postia leucomallella (Murrill) Jülich |
+ |
||||||
|
Postia sericeomollis (Romell) Jülich |
+ |
||||||
|
Postia undosa (Peck) Jülich |
+ |
||||||
|
Schizophyllum commune Fr.: Fr. |
+ |
+ |
+ |
+ |
+ |
||
|
Skeletocutis amorpha (Fr.) Kotl. & Pouzar |
+ |
||||||
|
Skeletocutis subincarnata (Peck) Keller |
+ |
+ |
|||||
|
Spongipellis spumeus (Sowerby: Fr.) Pat. |
+ |
+ |
|||||
|
Steccherinum aridum Svrcek |
+ |
||||||
|
Steccherinum fimbriatum (Pers.:Fr.) J.Erikss. |
+ |
+ |
|||||
|
Steccherinum nitidum (Pers.:Fr.) Vesterholt |
+ |
||||||
|
Steccherinum ochraceum (Fr.) Gray |
+ |
+ |
+ |
+ |
+ |
||
|
Stereum hirsutum hirsutum (Willd.: .) Gray |
+ |
+ |
+ |
+ |
|||
|
Stereum rugosum Pers.:Fr. |
+ |
||||||
|
Stereum subtomentosum Pouzar |
+ |
+ |
+ |
+ |
+ |
||
|
Thelephora terrestris Ehrh.: Fr. |
+ |
+ |
|||||
|
Tomentella cinerascens (P.Karst.) Hohn. & Litsch. |
+ |
||||||
|
Trametes gibbosa (Pers.: Fr.) Fr. |
+ |
+ |
|||||
|
Trametes hirsuta (Wulfen: Fr.) Pilat |
+ |
+ |
+ |
||||
|
Trametes ljubarskyi Pilat |
+ |
+ |
|||||
|
Trametes ochracea (Pers.) Gilb.& Ryvarden |
+ |
+ |
+ |
||||
|
Trametes pubescens (Schumach.: Fr.) Pilat |
+ |
+ |
+ |
+ |
|||
|
Trametes suaveolens (Fr.) Fr. |
+ |
+ |
|||||
|
Trametes Trogii Berk. |
+ |
+ |
+ |
||||
|
Trametes versicolor (L.: Fr.) Pilat |
+ |
+ |
+ |
+ |
+ |
||
|
Tremella mesenterica Retz.:Fr. |
+ |
+ |
+ |
||||
|
Trichaptum pargamenum (Fr.) G.Cunn. |
+ |
||||||
|
Trichaptum fuscoviolaceum (Ehrenb.: Fr.) Ryvarden |
+ |
||||||
|
Tubulicrinis borealis J.Erikss. |
+ |
||||||
|
Tubulicrinis propinquus (Bourdot & Galzin) Donk |
+ |
||||||
|
Volvariella bombycina (Schaeff.: Fr.) Singer |
+ |
||||||
Примечание: * Районы: 1 – Новосергиевский; 2 – Акбулакский; 3 – Оренбургский; 4 – Соль-Илецкий; 5 – Сакмарский; 6 – Илекский; 7 – Октябрьский.
Наибольшее разнообразие удалось выявить в Оренбургском, а также Новосергиевском районах. Количество обнаруженных видов варьировалось в очень широких пределах – от 16 в Илекском до 77 в Оренбургском районе. Столь значительные отличия в разнообразии определяются рядом причин. На первом месте – разный охват исследованиями территорий в разных районах, что обусловлено разной лесистостью. Также во многих районах (Оренбургский, Сакмарский, Илекский) изучались леса, произрастающие в поймах крупных рек (Урал, Сакмара) и занимающие значительные площади, в связи с чем их видовое разнообразие и богатство было заметно выше собственно степных лесов. К числу видов, связанных с пойменными лесами относятся, в частности, Inocutis dryophila, Fomitoporia robusta, Laetiporus sulphureus, Phellinus igniarius и др. Еще один немаловажный фактор – наличие в ряде районов (в особенности – в Новосергиевском районе) старовозрастных искусственных насаждений, в которых сформировалась специфичная фитосреда, обусловившая высокое разнообразие микокомплексов (в том числе и наличие ряда видов, не отмеченных в естественных древостоях того же видового состава) [1, 6]. К видам, специфичным для искусственных насаждений (в первую очередь – сосны) относятся, в частности, виды рода Postia.
Сравнение видового состава грибов посредством коэффициента сходства Съеренсена-Чекановского и последующий кластерный анализ позволил построить дендрит, наглядно показывающий взаимосвязи изученных микобиот (рис. 2).
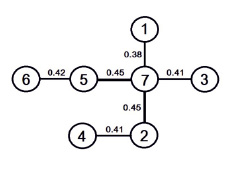
Рис. 2. Сходство видового состава древоразрушающих грибов районов исследований. Районы: 1 – Новосергиевский; 2 – Акбулакский; 3 – Оренбургский; 4 – Соль-Илецкий; 5 – Сакмарский; 6 – Илекский; 7 – Октябрьский
В целом, сходство между микобиотами достаточно низкое, и его максимум достигает 45 %. Вместе с тем, анализ существующих связей показывает более значимое сходство видового состава между микобиотами районов, расположенных непосредственно в пределах Оренбургского Предуралья, примерно на одной широте (Октябрьский, Сакмарский, Акбулакский районы). Микобиоты прочих районов, расположенных западнее и южнее, могут рассматриваться как производные от них. Если быть более точным, в районах, находящихся западней, выше доля видов, характерных для лесостепного микокомплекса [2, 4].
Таким образом, можно сделать вывод о типичности биоты ксилотрофных грибов центрального Оренбуржья для степной зоны региона. При этом имеется и ряд особенностей, отличающих изученную биоту от более западных степных районов. Многие обнаруженные нами виды широко распространены в регионе, но другие являются редкими. Ряд видов представлен единичными находками, поэтому их природоохранный статус пока трудно определить объективно, а другие официально отнесены к редким и исчезающим. Это, в частности, реликтовые виды, такие как Laetiporus sulphureus, Lenzites warnieri, Spongipellis spumeus, Trametes ljubarskyi [3, 7, 10].
Дальнейшее изучение биоты грибов-макромицетов центральных районов Оренбургской области является необходимым для объективной оценки биоресурсного потенциала региона.
Исследования выполнены при финансовой поддержке гранта губернатора и правительства Оренбургской области «Внедрение методов комплексной биолого-экологической и социально-экономической оценки биоресурсного потенциала центрального Оренбуржья как основы для формирования биотехнологического кластера региона» (2014 г.).
Рецензенты:
Русанов А.М., д.б.н., профессор, заведующий кафедрой общей биологии ФГБОУ ВПО «Оренбургский государственный университет», г. Оренбург;
Мирошников С.А., д.б.н., профессор, директор ГНУ ВНИИМС, г. Оренбург.
Работа поступила в редакцию 18.08.2014.
Библиографическая ссылка
Сафонов М.А., Маленкова А.С., Богомолова О.И., Русаков А.В. БИОРАЗНООБРАЗИЕ ДРЕВОРАЗРУШАЮЩИХ ГРИБОВ СТЕПНЫХ РАЙОНОВ ОРЕНБУРГСКОГО ПРЕДУРАЛЬЯ // Фундаментальные исследования. – 2014. – № 9-8. – С. 1750-1755;URL: https://fundamental-research.ru/ru/article/view?id=35134 (дата обращения: 25.04.2024).