



В системе защиты растений все более актуальным становится переход от борьбы с вредителями к созданию сбалансированных агроценотических систем. Массовые размножения многих видов вредных насекомых можно предотвратить или существенно снизить их вероятность при оптимизации структуры агроландшафта и пространственного расположения его элементов. Создание полимерного ландшафта с широким спектром различных антропогенных и естественных элементов может позволить добиться большей устойчивости агроэкосистем за счет увеличения численности ряда энтомофагов и стабилизации энтомокомплексов [1, 2, 3, 4].
Среди различных групп энтомофагов важное место отводится неспециализированным хищникам, численность которых не зависит от определенного вида жертвы. К таким энтомофагам относится большинство хищных напочвенных жесткокрылых, среди которых особый интерес вызывают представители жуков-жужелиц (Coleoptera, Carabidae), являющихся одной из основных групп почвенной мезофауны, зачастую доминируя как по численному, так и по видовому составу. Большой интерес жужелицы представляют не только как перспективные энтомофаги, подавляющие беспозвоночных вредителей на различных стадиях развития, но и как индикаторы экологических условий в естественных и антропогенных биотопах. Широкое использование видов этого семейства для биологического подавления вредителей в агроценозах затруднено. Неэффективность массового разведения хищных жужелиц в искусственных условиях для выпуска на посевы сельскохозяйственных культур, как это делается с некоторыми видами паразитических перепончатокрылых, приводит к тому, что основным направлением работы по использованию жужелиц в качестве энтомофагов является изучение возможностей увеличения их численности на посевах или сохранения их обилия при различных способах возделывания сельскохозяйственных культур, изучение сравнительного значения различных естественных биотопов, как мест размножения и резервации жужелиц и их роли, как источников формирования карабидокомплексов агроценозов. Выявление центров расселения жужелиц на посевы сельскохозяйственных культур позволит за счет их сохранения и, возможно, искусственного создания таких биотопов по соседству с полями постоянно поддерживать их высокое обилие в агроценозах. Однако до настоящего времени проведено недостаточно целенаправленных исследований, позволяющих сделать обоснованные выводы по указанным вопросам.
В орошаемых специализированных севооборотах, отличающихся высокой интенсификацией всех звеньев технологии производства сельскохозяйственной продукции, возрастает значение энтомофагов, так как использование химических методов в условиях орошаемого земледелия становится особенно опасным для окружающей среды. Наличие постоянных и временных оросителей, сети водосбросных каналов на орошаемых землях способствует распространению токсических препаратов с поливной водой на значительные площади, увеличивает вероятность их попадания в водоемы, используемые для водоснабжения населенных пунктов и животноводческих ферм, как водопои при выпасе. Поэтому именно для орошаемых массивов необходимо, в первую очередь, разрабатывать системы защиты урожая от вредителей, основанные на агротехнических, биологических и других безопасных методах, при минимальном использовании пестицидов. Все это указывает на острую необходимость тщательного изучения воздействия поливов на полезную энтомофауну агроценозов. Сведений по влиянию орошения на население жужелиц и других напочвенных жесткокрылых в условиях полупустыни недостаточно [5, 6, 10].
В настоящей статье представлены результаты изучения состава и структуры населения напочвенных жесткокрылых орошаемых агроландшафтов в условиях полупустыни Нижнего Поволжья.
Цель исследования
Выявить закономерности распределения и пути миграций жесткокрылых – энтомофагов напочвенного яруса агроэкосистем полупустынной зоны Нижнего Поволжья для оптимизации структуры орошаемых агроландшафтов и стабилизации их фитосанитарного состояния.
Материалы и методы исследования
Исследования проводились в 2010–2014 гг. на полях с орошением и прилегающих к ним биотопах на территории ФГУП «Орошаемое», располагающегося на границе степной и полупустынной зон в 20 км западнее г. Волгограда (Волгоградская область) (правобережье реки Волги). Почвы полей светло-каштановые в комплексе с солонцами.
Основной материал собран с помощью ловушек Барбера, в качестве которых использованы одноразовые пластиковые стаканы емкостью 0,2 л и диаметром отверстия 70 мм. Ловушки устанавливали по 10 штук, на расстоянии 10 м друг от друга. Выемка насекомых проводилась еженедельно, на полях – с начала вегетации до уборки культуры, в естественных местообитаниях – с апреля до октября.
Учитывая тот факт, что значительное число видов жесткокрылых на посевах сельскохозяйственных культур являются мигрантами из прилегающих местообитаний [7, 11], нами проводились синхронные учеты и наблюдения в различных типах биотопов, граничащих с севооборотным участком.
За время исследований в 21 биотопе было собрано и определено свыше 65 тысяч экземпляров отряда Coleoptera. Динамическая плотность особей выражена в числе экземпляров на 10 ловушко-суток (в тексте экз./10 лов.-сут.). Всего отработано 57 тыс. л.-сут. Анализ сообществ проводили по представителям 4 семейств – жужелиц (Carabidae), чернотелок (Tenebrionidae), кожеедов (Dermestidae) и быстрянок (Anthicidae). Структура населения герпетобионтов характеризовалась по нескольким экологическим параметрам: видовому составу, численности в единицах динамической плотности, структуре доминирования, спектрам биотопических групп и жизненных форм и сезонной динамике активности.
Структура доминирования определялась по следующим критериям: доминанты – 5 и более %, субдоминанты 2–5 %, редкие – менее 2 % .
Для сравнения комплексов жесткокрылых в различных биотопах и установления взаимосвязей между ними проводили анализ на основе мер сходства. Их расчет вели по формуле Чекановского-Соренсена, модифицированной для учета сравнительного обилия видов [8, 12].
Результаты исследования и их обсуждение
В районе исследований на ограниченном участке орошаемого агроландшафта ловушками за 5 лет учтено 136 видов из рассматриваемых 4 семейств жесткокрылых. Жужелицы являлись в напочвенном ярусе абсолютными доминантами по числу видов, а на полях – и по относительной численности особей. Особенности распределения представителей отдельных семейств по элементам агроландшафта представлены в табл. 1.
Рассматривая структуру комплексов напочвенных жесткокрылых по элементам агроландшафта (табл. 2), следует отметить, что по доле жужелиц в общей структуре комплекса можно выделить две группы полей. Первая включает посевы многолетних трав и озимой пшеницы (50–67 %), а вторая – пропашные культуры (89 % и более).
В табл. 3 представлены данные о видовом составе основного ядра комплексов жесткокрылых на полях и роли в них отдельных видов. За исключением полей озимой пшеницы и люцерны второго года жизни, на всех остальных полях максимальная численность (суммарно за сезон) у жужелиц отмечена для жужелицы волосистой (Harpalus rufipes), у чернотелок доминировал медляк песчаный малый (Gonocepalum pusillum), у быстрянок массовыми были Hirticomus hispidus и Anthicus tristis, а среди кожеедов – Dermestes coronatus.
Жужелица волосистая (H. rufipes), по системе жизненных форм И.Х. Шаровой [9], относится к миксофитофагам с преобладающей фитофагией. В 50–60-е годы прошлого века многие авторы считали этот вид вредителем зерновых, однако исследования последних десятилетий показывают, что он может играть существенную роль в регулировании численности различных беспозвоночных-фитофагов. Объектами питания жужелицы волосистой являются имаго и личинки вредной черепашки, проволочники, личинки колорадского жука и др. [4, 5].
Таблица 1
Видовое разнообразие и численность напочвенных жесткокрылых в различных биотопах агроландшафта
|
№ п/п |
Биотопы |
Жужелицы |
Чернотелки |
Быстрянки |
Кожееды |
|
1 |
Люцерна, второй год жизни |
45/20,3* |
6/7,2 |
5/2,7 |
3/3,7 |
|
2 |
То же, третий |
43/16,2 |
6/4,6 |
6/3,4 |
4/4,9 |
|
3 |
То же, четвертый |
42/32,6 |
7/4,4 |
6/3,2 |
3/4,7 |
|
4. |
Озимая пшеница |
42/15,6 |
5/3,8 |
5/2,7 |
3/7,1 |
|
5 |
Кукуруза на зерно |
25/3,0 |
2/0,05 |
3/0,03 |
1/0,01 |
|
6 |
Свекла кормовая |
29/18,3 |
2/0,2 |
4/0,8 |
2/0,08 |
|
7 |
Картофель |
26/7,9 |
2/0,1 |
2/0,6 |
2/0,03 |
|
8 |
Лесополоса, вяз |
31/2,2 |
8/2,1 |
5/0,4 |
4/0,1 |
|
9 |
Целинная степь, плакор |
29/5,7 |
11/5,8 |
3/0,1 |
1/0,04 |
|
10 |
Балка, склон, северной экспозиции |
44/3,6 |
11/4,3 |
4/0,5 |
2/0,3 |
|
11 |
То же, южной экспозиции |
25/2,6 |
12/5,3 |
2/0,02 |
1/0,05 |
|
12 |
Балка, дно |
41/2,5 |
5/2,5 |
6/0,8 |
4/2,9 |
Примечание. * в числителе – число видов, в знаменателе – численность в экз./10 л.-с.
Таблица 2
Структура комплексов напочвенных жесткокрылых на полях с посевами различных культур (в % от общей численности)
|
№ п/п |
Биотопы |
Жужелицы |
Чернотелки |
Быстрянки |
Кожееды |
Прочие |
|
1 |
Люцерна, второй год жизни |
58,1 |
20,6 |
7,7 |
10,6 |
2,8 |
|
2 |
То же, третий |
53,2 |
15,1 |
11,2 |
16,1 |
4,4 |
|
3 |
То же, четвертый |
66,7 |
9,0 |
6,5 |
9,6 |
8,1 |
|
4 |
Озимая пшеница |
50,1 |
12,2 |
8,7 |
22,8 |
6,2 |
|
5 |
Кукуруза на зерно |
95,7 |
1,6 |
1,0 |
0,3 |
1,4 |
|
6 |
Свекла кормовая |
92,3 |
1,0 |
4,0 |
0,5 |
2,2 |
|
7 |
Картофель |
89,0 |
1,1 |
6,8 |
0,4 |
2,7 |
Таблица 3
Доля участия массовых видов жесткокрылых в структуре энтомокомплексов герпетобия различных культур
|
Виды жесткокрылых |
% от общей численности жесткокрылых |
||||||
|
1* |
2 |
3 |
4 |
5 |
6 |
7 |
|
|
Carabidae |
|||||||
|
Calosoma auropunctatum |
3,0 |
10,4 |
11,8 |
2,6 |
0,9 |
1,1 |
0,7 |
|
C. denticolle |
3,0 |
4,6 |
2,5 |
2,0 |
0,2 |
0,6 |
0,2 |
|
Poecilus cupreus |
20,0 |
4,1 |
5,2 |
15,5 |
0,5 |
5,1 |
3,0 |
|
P. nitens |
2,6 |
2,0 |
1,5 |
4,5 |
0,5 |
0,0 |
1,8 |
|
P. puncticollis |
2,7 |
2,1 |
3,9 |
6,0 |
0,6 |
0,7 |
2,7 |
|
Harpalus rufipes |
6,6 |
13,3 |
31,6 |
3,2 |
76,6 |
53,3 |
63,2 |
|
H. distinguendus |
2,0 |
4,0 |
1,5 |
0,9 |
0,9 |
2,9 |
5,3 |
|
Прочие |
18,2 |
12,7 |
8,7 |
14,8 |
15,5 |
28,4 |
12,1 |
|
Tenebrionidae |
|||||||
|
Gonocephalum pusillum |
19,8 |
13,1 |
7,6 |
12,1 |
1,55 |
0,93 |
1,0 |
|
Прочие |
0,8 |
2,0 |
1,4 |
0,1 |
0,05 |
0,07 |
0,1 |
|
Anthicidae |
|||||||
|
Anthicus tristis |
2,3 |
5,4 |
2,2 |
5,2 |
0,09 |
0,68 |
0,8 |
|
A. subcrucifer |
1,2 |
0,6 |
0,5 |
1,0 |
0,01 |
0,06 |
0.2 |
|
Hirticomus hispidus |
3,1 |
3,9 |
2,6 |
1,8 |
0,90 |
3,16 |
5,6 |
|
Прочие |
1,1 |
1,3 |
1,2 |
0,2 |
0,00 |
0,10 |
0,2 |
|
Dermestidae |
|||||||
|
Dermestes coronatus |
8,8 |
11,3 |
7,1 |
19,1 |
0,3 |
0,47 |
0,36 |
|
D. kaszabi |
1,7 |
3,9 |
2,4 |
2,9 |
0,0 |
0,00 |
0,00 |
|
Прочие |
0,1 |
0,9 |
0,1 |
0,8 |
0,0 |
0,00 |
0,00 |
Примечание. * нумерация полей как в табл. 2.
По данным наших исследований, жужелица волосистая на люцерновых полях отдает явное предпочтение животной пище. Только у 14 % из 176 вскрытых жуков в желудке обнаружены остатки растительной пищи, тогда как у 73 % содержалось большое количество остатков хитина. В отдельных случаях по остаткам удалось идентифицировать вид жертвы. Установлено, что около 35 % жуков питались гусеницами лугового мотылька и люцерновой совки. Отмечены единичные случаи питания жуками тихиуса желтого (Tichius flavus).
Близость полей люцерны сказывается на численности жужелиц на участках с пропашными культурами. На половине поля, граничащего с люцерной, численность этих хищников увеличивается в 1,6–2,7 раза в сравнении с противоположной половиной, частично изолированной от многолетних трав оросителем.
Анализ взаимосвязей между комплексами жесткокрылых различных участков агроландшафта на основе мер сходства (рисунок) показал, что их можно разделить на несколько естественных групп. К первой относятся группировки жесткокрылых аридных естественных местообитаний, ко второй – посевов озимой пшеницы, люцерны и мезофитного лугового разнотравья на дне балки, к третьей – полей пропашных культур. Комплекс жесткокрылых лесополосы занимает промежуточное положение между группировками аридных степных биотопов и полями.
Условные обозначения:
1 – люцерна 2 года жизни,
2 – люцерна 3 года жизни,
3 – люцерна 4 года жизни,
4 – озимая пшеница,
5 – кукуруза,
6 – свекла,
7 – картофель,
8 – лесополоса,
9 – целинная степь, плакор,
10 – склон балки N экспозиции,
11 – склон балки S экспозиции,
12 – дно балки.
Изучение напочвенных жесткокрылых в агроландшафте, включающем орошаемые поля с посевами различных культур, лесополосы и сохранившиеся естественные местообитания, показало, что среди этой группы энтомофауны преобладают многоядные хищники семейства жужелиц. При этом на площади около 50 га только с помощью почвенных ловушек за 5 лет отмечено более четверти суммарного видового разнообразия жужелиц в регионе.
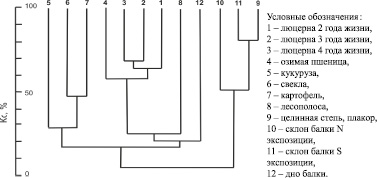
Дендрограмма сходства комплексов жесткокрылых различных элементов агроландшафта по коэффициенту сходства с учетом обилия общих видов (Кс)
В естественных биотопах, особенно ксерофитных, доля жужелиц сравнительно невелика, тогда как на полях достигает 90 % и более. На посевах доминируют виды, характерные для агроценозов практически на всей территории европейской части СССР (Poecilus cupreus, Harpalus rufipes, H. distinguendus), полей степной зоны (Calosoma auropunctatum, C. denticolle), а также типичные для зоны сухих степей и полупустынь Юго-Востока, галофильные Poecilus puncticollis и P. nitens.
Анализ литературных данных показывает, что массовые виды жужелиц района исследований являются регуляторами численности клопа-черепашки, пшеничного трипса, проволочников, колорадского жука, гусениц серой зерновой совки и др. Данные по специфике питания жужелицы волосистой, полученные нами, позволяют, учитывая высокую численность этого вида, считать её, наряду с красотелами, важным регулятором численности вредных чешуекрылых [5].
На полях с различными культурами складываются неравноценные условия для обитания напочвенных жесткокрылых. Различия обусловлены особенностями микроклимата, создающегося в напочвенном ярусе растениями, а также фенологическими особенностями культуры и технологией её возделывания. На посевах культур, технологический цикл выращивания которых от посева до уборки ограничен несколькими месяцами весенне-летнего периода, жужелицы являются временными обитателями. Это обусловлено ежегодными «катастрофическими» воздействиями на экосистему поля, не позволяющими абсолютному большинству видов проходить здесь цикл развития. Посевы озимой пшеницы более благоприятны в этом отношении, однако, как показывают наблюдения, уборка и последующая обработка почвы в начале июля совпадают с периодом максимального обилия и активности личинок красотелов и ведут к их массовой гибели.
Оптимальные условия для обитания, размножения и развития полевых жужелиц создаются на полях многолетних трав, в первую очередь, люцерны. Возделывание её на одном поле 3–4 года, благоприятный микроклимат напочвенного яруса и высокая численность беспозвоночных определяют значительное видовое разнообразие и высокое обилие жужелиц. Здесь четко выражены два максимума численности и активности видов этого семейства в течение сезона. Первый (май-июнь) формируют преимущественно виды родов Calosoma и Poecilus, а второй (июль-август) создается практически за счет одного вида – H. rufipes. Кроме указанных, в конце августа – сентябре наблюдается третий пик активности за счет появления молодых имаго видов весенне-летней фенологической группы: Poecilus cupreus, P. puncticollis, P. crenuliger, P. nitens.
Наиболее неблагоприятные условия для жужелиц складываются на пропашных культурах поздних сроков сева. Такие поля в течение длительного периода (до середины июля и более) подвергаются интенсивным механическим и химическим (гербициды) воздействиям; растительный покров, формирующий необходимый для жужелиц микроклимат, образуется также лишь в начале лета. Эти причины определяют низкое видовое разнообразие энтомокомплексов герпетобия полей пропашных культур. Более 90 % жесткокрылых здесь составляют жужелицы, мигрирующие на поля из прилегающих местообитаний с середины июня, при этом свыше 70 % их численности образуется за счет жужелицы волосистой. Низкое видовое разнообразие и монодоминантная структура комплексов жесткокрылых на полях пропашных культур указывают на неустойчивость сообщества, формирующегося на этих участках агроландшафта.
В качестве единого агроценоза мы рассматриваем участок агроландшафта, включающий поля с различными культурами и прилегающие к ним естественные и антропогенные биотопы. Это справедливо, в первую очередь, для орошаемого земледелия, где площадь отдельных полей в севообороте составляет от 5–10 до 50–70 га. В таких условиях возможно обогащение энтомокомплексов пропашных культур за счет миграций энтомофагов. Источниками таких миграций могут быть участки естественного разнотравья, полезащитные лесополосы и люцерновые поля, комплексы жесткокрылых в которых наиболее близки по коэффициенту сходства к комплексам пропашных культур.
Оценивая сравнительное значение различных местообитаний как возможных источников увеличения численности жужелиц за счет миграций, следует отметить, что сравнительная численность этих энтомофагов в естественных биотопах и лесополосе значительно меньше, чем на полях. Учитывая незначительную площадь, занимаемую такими биотопами, нельзя считать возможной миграцию из них значительного числа особей жужелиц. Кроме того, массовыми видами здесь являются практически не встречающиеся на полях Carabus hungaricus, C. estreicheri, Calathus distinguendus, Pterostichus niger, Ophonus azureus и др. Такие элементы агроландшафта могут служить для полей источниками редких видов и местами зимовки и размножения жужелиц [7], но не способны существенно увеличить их обилие на посевах за счет миграций.
Эффективными источниками массовых миграций жужелиц в районе исследований являются многолетние травы. На посевах пропашных культур, граничащих с полями люцерны, на 60–170 % увеличивается относительная численность этих энтомофагов. Аналогичные данные получены в Ростовской обл., где 40 %-ое увеличение численности жужелиц зарегистрировано в 150 м от границы с посевом люцерны [11].
Заключение
Таким образом, можно сделать заключение, что для увеличения численности и равномерного распределения хищных напочвенных жесткокрылых в севообороте необходимо наличие в его структуре многолетних трав, поля которых должны располагаться вблизи участков с наиболее интенсивной антропогенной нагрузкой. Соблюдение таких условий позволит обеспечивать высокую численность и увеличивать разнообразие энтомофагов на посевах пропашных культур за счет горизонтальных миграций, а в результате будет способствовать большей стабильности энтомокомплекса агроценоза в целом.
Учитывая важное значение люцерновых полей как мест накопления, размножения и сохранения напочвенных многоядных хищников, следует предусматривать минимальное использование пестицидов на семенниках этой культуры.
Рецензенты:
Иванцова Е.А., д.с.-х.н., доцент, заведующая кафедрой экологии и природопользования ФГАОУ ВПО Волгоградского государственного университета, г. Волгоград;
Москвичев А.Ю., д.с.-х.н., профессор кафедры агроэкологии и защиты растений ФГБОУ ВПО Волгоградского государственного аграрного университета, г. Волгоград.
Работа поступила в редакцию 30.12.2014.
Библиографическая ссылка
Комаров Е.В., Карпова Т.Л. СОСТАВ, СТРУКТУРА, РАСПРЕДЕЛЕНИЕ КОМПЛЕКСОВ ГЕРПЕТОБИОНТНЫХ ЖЕСТКОКРЫЛЫХ (COLEOPTERA) В ОРОШАЕМОМ АГРОЛАНДШАФТЕ // Фундаментальные исследования. – 2014. – № 12-11. – С. 2350-2356;URL: https://fundamental-research.ru/ru/article/view?id=36694 (дата обращения: 25.04.2024).