



Перифитонные сообщества занимают важное место в формировании структуры и функционировании водных экосистем. Это определяется как таксономическим и функциональным многообразием, так и большой активностью метаболических процессов, высокими показателями обилия перифитонных организмов [7]. Сообщества обрастателей, формирующиеся на границе раздела фаз твердый субстрат – вода, играют ключевую роль в организации пространственной структуры экосистем. Эти биотопы всегда характеризуются высоким разнообразием и повышенными количественными показателями в силу действия пограничного эффекта. Выраженность данного эффекта зависит от сложности архитектоники субстрата, на котором формируется сообщество. Это понятие включает элементы, составляющие поверхность, их соотношение и взаиморасположение, образующее топологию субстрата. Архитектоника изменяется также в зависимости от расположения в пространстве и взаимодействия близлежащих поверхностей. Таким образом, жизненное пространство перифитона имеет мозаичную и иерархическую организацию, что отражается на особенностях формирования экологической структуры прикрепленных сообществ. В то же время возможность заселения субстрата разными видами зависит от их экологии и морфологических адаптаций, связанных с прикреплением к поверхности.
Актуальность изучения процессов колонизации погруженных поверхностей увеличивается в связи с проблемой захламленности водных объектов и, как следствие, появления в них новых субстратов антропогенного происхождения. Удобной модельной группой для исследований процесса колонизации любого субстрата являются перифитонные инфузории. Высокая скорость размножения инфузорий и быстрое чередование поколений позволяет в относительно короткие сроки получить данные о динамике популяций и видовых комплексов [4]. Небольшие размеры и компактность микросообществ предоставляют также широкие возможности для эксперимента с их формированием на разнотипных субстратах.
Цель работы – проанализировать формирование экологической структуры сообществ через освоение субстратов разной архитектоники экоморфологическими группами прикрепленных инфузорий.
Материалы и методы исследования
Изучались сообщества инфузорий, формирующиеся на модельных (стекла обрастания) и естественных субстратах в разнотипных водоёмах Вологодской области. Этот набор включал небольшой водоток, среднюю и крупную реку и водохранилище. С одной стороны, это давало возможность учесть значение разномасштабности микросообществ и водных объектов, а с другой – влияние течения на формирование цилиоперифитонных сообществ.
К небольшим водным объектам, в которых проводились сборы обрастаний, относятся мелиоративная канава в окрестностях города Вологды и речка Большой Пучкас. Средняя и крупная реки представлены Вологдой и Сухоной соответственно, крупный водный объект озерного типа – Рыбинским водохранилищем. Все исследованные водотоки являются взаимосвязанной системой, так как мелиоративная канава соединяется с рекой Вологдой, которая, в свою очередь, как и река Большой Пучкас, является притоком Сухоны.
Сборы проводились по стандартным методикам в 2006–2013 годах [3]. Инфузории наблюдались непосредственно на стёклах обрастания, или в соскобах с поверхности субстрата прижизненно при увеличении микроскопа 10×15 и 10×20. Для определения видов использовались общепринятые определители и таблицы [2, 6, 8]. Биомассу клеток определяли объёмным методом, уподобляя её геометрической фигуре и измеряя их линейные размеры; плотность полагали равной единице [3]. Для сравнительного анализа структуры прикрепленных сообществ на разных субстратах изучался видовой состав, численность и биомасса инфузорий. Статистическая обработка результатов осуществлялась с помощью пакета «GRAPHS».
Результаты исследования и их обсуждение
Результаты исследований подтвердили, что характер морфологических адаптаций сидячих инфузорий накладывает отпечаток на особенности освоения ими пространства и позволяет уменьшить топическую конкуренцию между экологическими группами. Исходя из этого, проанализированы основные адаптации к прикрепленному образу жизни. Известно, что для инфузорий характерны разные способы прикрепления, выделяются стебельковые и бесстебельковые формы. Последние прикрепляются к субстрату своеобразной подошвой, напоминающей присоску, некоторые из них способны передвигаться, шагая по субстрату. В исследуемых водоёмах видовое разнообразие бесстебельковых инфузорий невелико, на них приходится 9–18 % от общего числа видов водотока. Такое соотношение видового богатства типично для разнотипных водных объектов, в том числе и по известным литературным данным [1, 5].
Бесстебельковые инфузории характеризуются низкой эффективностью использования поверхности субстрата, доступной для прикрепления, поскольку подошва занимает некоторую площадь. Кроме того, они не могут эффективно участвовать в реализации ярусности перифитона. Громоздкое, по сравнению со стебельком других форм, тело ограничивает подвижность данных инфузорий в пространстве и препятствует прикреплению рядом других сидячих видов. Это подтверждается низкими значениями численности этих инфузорий на различных субстратах (рис. 1).
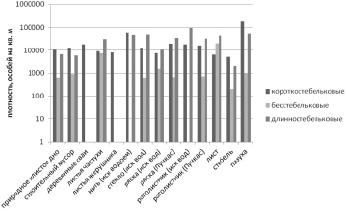
Рис. 1. Численность представителей различных экоморфологических групп инфузорий на различных субстратах (построенная в логарифмической шкале)
В тоже время конкурентоспособность бесстебельковых форм в освоении пространства повышается, во-первых, за счет возможности перемещения особей. Это дает некоторое преимущество для освоения новых участков по сравнению неподвижными стебельковыми формами. Во-вторых, значительные размеры большинства представителей этой группы обуславливают их значительный вклад в биомассу прикрепленных сообществ, на простых субстратах превышающий 90 %. В-третьих, высокая подвижность тела инфузории, обуславливающая разнообразие положений цитостома в пространстве, способствует эффективности питания.
Благодаря этим преимуществам бесстебельковые инфузории способны эффективно вытеснять из сообщества виды других экологических форм в конкуренции за субстрат. Увеличение численности этой группы видов сопровождается ростом их вклада в общую численность исследованных сообществ. Доля сидячих инфузорий в суммарной численности цилиоперифитонного сообщества достоверно коррелирует с их численностью. Эта закономерность подтверждается сравнительным анализом структуры сообществ различных микроместообитаний, включая разные типы естественных и искусственных субстратов.
Представленность сидячих видов более вариабельна в сообществах, формирующихся на разнотипных по архитектонике субстратах, чем на однотипных в различных водотоках. Различие показателей представленности бесстебельковых видов не связано с общим видовым богатством сообществ, формирующихся на данном субстрате: достоверность аппроксимации этих показателей составляет лишь 0,03.
Большая часть видов в исследуемых сообществах представлена стебельковыми формами (80–90 %), которые более эффективно осваивают поверхность субстрата за счет особенностей морфологии. Стебелёк позволяет инфузориям выходить за пределы пограничного слоя, что обеспечивает им преимущество в снабжении пищей, приносимой течением. Подошва прикрепленной инфузории образует несократимый (триба Acontractilia) или сократимый (триба Contractilia) стебелек.
Результаты исследований выявили большее разнообразие в разных сообществах представителей трибы Сontractilia, которые составляют от 23 до 42 % видового разнообразия, что значительно больше показателей представителей трибы Acontractilia (доля которых в числе видов не превышает 18 %). В разнотипных водоемах эти показатели несколько отличаются, однако корреляции с типом водоема или степенью его изученности не выявлено.
Длинно-стебельковые виды преобладают в цилиоперифитонных сообществах и вносят наиболее значительный вклад в их численность. Многочисленность длинно-стебельковых форм не всегда соответствует высокой доле в общей численности перифитонного сообщества. Отсутствие выраженной зависимости между представленностью этих форм в сообществе и её абсолютным значением их численности объясняется существованием нескольких механизмов снижения конкуренции. С одной стороны, в двухмерном пространстве они конкурируют за место прикрепления с крупными сидячими особями. С другой стороны, благодаря наличию стебелька, такие инфузории характеризуются очень маленькой площадью прикрепления. Значительная, по сравнению с размером клетки длина стебелька, позволяет формировать сложную кустовую структуру и размещать зоиды в трехмерном пространстве.
На формы, обладающие небольшим несократимым стебельком, приходится в среднем 50 % видов инфузорий перифитонного сообщества. Они не получают преимущества, связанного с выносом перистома из слоя вытеснения. Такие инфузории, как правило, имеют небольшие размеры, перемещение цитостома в пространстве может достигаться лишь через сокращение или растяжение тела. Помимо того домик также в значительно ограничивает возможности изменения положения цитостома в пространстве.
Отсутствие мобильности у коротко-стебельковых форм компенсируется широкими возможностями прикрепления к различным субстратам. Неподвижность и малая длина стебелька позволяют им прикрепляться в местах, недоступных для крупных или длинно-стебельковых инфузорий, например, к поверхности диатомовых или даже стебелькам крупных или колониальных инфузорий. Таким образом, с одной стороны, формируется ярусная структура перифитонного сообщества, а с другой – может ослабляться конкуренция за субстрат с другими формами прикрепленных инфузорий.
Доля коротко-стебельковых особей в общей численности перифитона не коррелирует с их абсолютной численностью. Суммарная плотность этих видов составляет в одинаковых условиях равную величину от 7 до 20 тысяч особей на квадратный метр. Таким образом, коротко-стебельковые формы достигают максимально возможной в данных условиях численности вне зависимости от их доминирования в сообществе. Это позволяет заключить, что они почти не испытывают конкурентного давления со стороны более крупных форм.
Конкурентоспособность вида в освоении субстрата повышается благодаря формированию колоний. Поэтому колониальность часто встречается у прикрепленных инфузорий в условиях обильного питания, например, в системах очистки сточных вод и на поверхности зоопланктона. Значительно более редкими являются находки колоний на погруженной растительности и других субстратах. Колонии инфузорий обнаружены лишь в пробах, взятых в условиях экстремальной сапробности, в реке Сухоне в подледный период, в месте сброса сточных вод.
Размер особи имеет большое значение для освоения пространства инфузориями, тем более что среди них встречаются виды, значительно различающиеся по размерам клетки. От размера особи зависит колонизация сложноорганизованного субстрата, на поверхности которого создаются пространственные ниши, характеризующиеся ограниченностью пространства, недоступные для крупных видов.
Наблюдается увеличение доли мелких видов в сообществе от общей численности в ряду микроместообитаний по степени усложнения структуры (рис. 2). Листья частухи представляют для инфузорий обширную поверхность простой топологии. Другие субстраты (илистое дно, строительный мусор, деревянные сваи) покрыты нитчатыми водорослями, которые формируют сложную трехмерную структуру различной плотности. Наибольшая плотность расположения талломов характерна для водорослей, покрывающих сваи, где они образуют плотный войлокообразный покров.
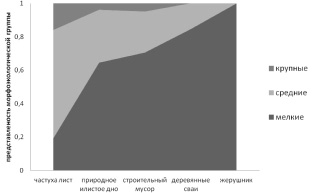
Рис. 2. Численность и доля разноразмерных видов в сообществах, формирующихся на различных субстратах в ряду усложнения их пространственной структуры
Самая сложная из рассмотренных субстратов архитектоника характерна для подводных сильнорассеченных листьев жерушника, местами также обильно покрытых нитчатыми водорослями. Отметим, что данный субстрат характеризуется одновременно низким разнообразием и невысокой численностью сообщества, что связано с трудностью его освоения (таблица).
Показатели видового разнообразия сообществ обрастаний на разнотипных субстратах в р. Вологда
|
Субстрат |
Деревянные сваи |
Строительный мусор |
Природное илистое дно |
Листья частухи |
Листья жерушника |
|
Значение индекса Шеннона для перифитонного сообщества |
3,31 |
3,12 |
2,81 |
1,95 |
1,33 |
Заключение
Анализ экологической структуры прикрепленных сообществ на разнотипных субстратах, как естественных, так и антропогенно привнесенных показал, что их освоение экоморфологическими группами инфузорий зависит от сложности архитектоники. Самый сложноорганизованный из рассмотренных субстратов – листья жерушника характеризуется наименьшим видовым разнообразием цилиосообществ. Наибольшее видовое разнообразие сообществ характерно для субстратов, занимающих промежуточные позиции в ряду сложности их архитектоники. Это нетипичные для водных экосистем субстраты антропогенного происхождения – деревянные сваи и строительный мусор. Это обусловлено разнообразием условий на их поверхности и формированием мозаичной структуры первичных обрастаний, представленных нитчатыми водорослями.
Показано, что формирование структуры сообществ связано с различной конкурентоспособностью экоморфологических групп инфузорий к освоению субстрата. Размер особи имеет ключевое значение для успешного освоения инфузориями пространственных ниш, причем на простом субстрате преимущество получают крупные формы, а на сложноорганизованном – мелкие.
Прикрепление к субстрату с помощью стебелька позволяет значительно снижать топическую конкуренцию, благодаря чему стебельковые инфузории характеризуются высокими значениями численности и более эффективно осваивают поверхность субстрата за счет создания ярусности. Крупностебельковые формы доминируют на всех субстратах, за исключением природного субстрата с самой сложной архитектоникой (листья жерушника). Более выражено их доминирование на простоорганизованных субстратах.
Формирование колоний свойственно лишь отдельным видам, на которые приходится от 10 до 70 % от видового состава, в некоторых микроместообитаниях колониальные виды не представлены. Установлено, что колониальные инфузории, для которых характерен крупный размер зоида, приурочены к пространственно-простым субстратам, что выражается в показателях относительной численности. Напротив, доля колониальных видов с мелким зоидом в общей численности выше на сложноорганизованных субстратах.
Таким образом, успешность освоения субстрата разными экоморфологическими группами определяется уровнем сложности организации его поверхности.
Рецензенты:
Болотова Н.Л., д.б.н., профессор, зав. кафедрой зоологии и экологии, ФГБОУ «Вологодский государственный педагогический университет», г. Вологда;
Мыльников А.П., д.б.н., главный научный сотрудник, ФГБУН «Институт биологии внутренних вод им. И.Д. Папанина» РАН, п. Борок, Ярославская обл.
Работа поступила в редакцию 05.12.2013.
Библиографическая ссылка
Мухин И.А. ОСВОЕНИЕ СУБСТРАТОВ РАЗНОЙ АРХИТЕКТОНИКИ ЭКОМОРФОЛОГИЧЕСКИМИ ГРУППАМИ ПРИКРЕПЛЕННЫХ ИНФУЗОРИЙ // Фундаментальные исследования. – 2013. – № 10-14. – С. 3115-3119;URL: https://fundamental-research.ru/ru/article/view?id=32971 (дата обращения: 26.04.2024).