



С точки зрения популяционного подхода экосистема рассматривается как сочетание популяций разных видов, относящихся к определенным трофическим группам и взаимодействующих между собой на различных уровнях [5, 6]. Основным условием устойчивого существования экосистем является наличие элементарных популяций – множества особей одного вида, необходимого и достаточного для устойчивого потока поколений в минимально возможном пространстве [7]. В лесных экосистемах основными эдификаторами являются древесные растения, поскольку именно они формируют мозаику светового, водного и почвенного режимов и тем самым оказывают существенное влияние на возможный набор «подчиненных» видов растений, животных, грибов [7]. В условиях юга российского Дальнего Востока наиболее мощными эдификаторами хвойно-широколиственных лесов являются Pinus koraiensis, Picea ajanensis, Abies nephrolepis, Betula costata, Tilia amurensis и ряд других видов.
Согласно теории gap-динамики, непрерывное существование лесных экосистем обеспечивается за счет процессов спорадического образования и зарастания «окон» в пологе древостоя. Считается, что зарастание окон происходит главным образом за счет подроста, появившегося в окне после его образования, в то время как подрост, существующий под пологом древостоя, выполняет роль буфера популяции [6]. В процессе изучения структуры и динамики кедрово-широколиственных лесов нами было установлено, что
а) подрост хвойных и лиственных видов образует под пологом древостоя скопления (группы);
б) размещение этих групп не имеет явной корреляции с расположением и конфигурацией окон в пологе;
в) развитие подроста хвойных видов протекает ступенчато, когда периоды быстрого роста чередуются с периодами угнетения [4, 9, 10].
Исходя из этого, было высказано предположение, что наблюдаемые группы подроста представляют собой результат длительного его накопления под пологом, и новое поколение может образоваться из подроста, имеющего значительно различный абсолютный возраст за счет одновременного перехода к одному и тому же онтогенетическому возрасту при улучшении условий. Цель данного исследования заключалась в выяснении абсолютного возраста подроста в группах (образуются ли группы за счет одновременного приживания подроста или это действительно длительное накопление), диапазона вариаций возраста подроста разных видов и возможных соотношений абсолютного и онтогенетического возраста.
Материалы и методы исследования
Исследование проведено на территории Верхнеуссурийского стационара Биолого-почвенного института Дальневосточного отделения РАН, расположенного на восточном макросклоне южной части горной системы Сихотэ-Алинь, в бассейне р. Правая Соколовка, на одной постоянной и трех временных пробных площадях. Постоянная пробная площадь (ППП) № 71-2012 размером 1,4 га (150×90 м) заложена в пределах участка коренных спелых лесов, относящихся к типичным кедровым лесам на верхней границе распространения кедра корейского, где он формирует смешанные формации кедрово-еловых и елово-широколиственных лесов. В окрестностях ППП были подобраны три участка с группами подроста, аналогичные тем, которые были описаны на данной постоянной пробной площади [4, 9, 10]. На этих участках были заложены три временные пробные площади (ВПП) размером 0,04 га (20×20 м) для изучения абсолютного возраста подроста хвойных видов и онтогенетической структуры групп. Вокруг пробных площадей был добавлен «буфер» шириной 5 м с каждой стороны для учета деревьев, стволы которых начинаются за пределами ВПП, но их кроны участвуют в образовании полога.
Древостой на постоянной и временных пробных площадях образован деревьями следующих видов: Abies nephrolepis (Trautv.) Maxim., Acer ukurunduense Trautv. et C.A. Mey., Picea ajanensis (Lindl. et Gord.) Fisch. ex Carr., Pinus koraiensis Siebold. еt Zucc., Tilia amurensis Rupr., Betula costata (Trautv.) Regel, единично – Taxus cuspidata Siebold & Zucc., Ulmus laciniata (Trautv.) Mayr, Sorbus pochuashanensis (Hance) Hedl., Acer mono Maxim. и Padus maximowiczii (Rupr.) Sokolov. Подлесок и лианы: Actinidia kolomikta Maxim. et Rupr., Schisandra chinensis (Turcz.) Baill., Ribes maximoviczianum Kom., Philadelphus tenuifolius Rupr. et Maxim., Acer barbinerve Maxim.
В древостое выделяются три подъяруса, включая подрост. В первом (верхнем) подъярусе преобладают деревья P. koraiensis, A. nephrolepis, P. ajanensis и T. amurensis, и, в меньшей степени B. costata. Второй подъярус образуют генеративные деревья A. ukurunduense и виргинильные деревья других видов. Они равномерно распределены по высоте подъяруса, многие из них находятся в угнетенном состоянии (крона деревьев P. ajanensis и A. nephrolepis имеет характерную «зонтиковидную» форму [8]. Подрост многочисленный, преобладают растения хвойных видов. Развитие крупного подроста (виргинильных растений) замедлено.
Ревизии постоянной пробной площади проведены в 2009–2012 гг. Во время ревизий собраны данные о древостое, включающие вид дерева, его относительные координаты, высоту, высоту начала кроны, диаметр дерева на высоте 1,3 м и у корневой шейки (для подроста ниже 1,3 м), проекции крон, онтогенетическое и жизненное состояния. Для учета подлеска и травяного яруса пробная площадь была подразделена на квадраты размером 2×2 метра, затем для каждого такого квадрата составлялся список видов кустарников, лиан и трав с указанием их проективного покрытия.
Работа на ВПП проведена в 2012 г. На трех ВПП были собраны данные по древостою, подлеску и травяному ярусу по методике, аналогичной для ППП. Помимо этого с каждой пробной площади отбирались образцы растений P. koraiensis, P. ajanensis и A. nephrolepis для определения абсолютного возраста подроста. В рамках этого исследования к подросту были отнесены растения в следующих возрастных состояниях: ювенильное (juv), имматурное в трех подгруппах (im1, im2, im3) и виргинильное первой подгруппы (v1). Разделение возрастных состояний P. koraiensis, P. ajanensis, A. nephrolepis проведено согласно описаниям, данным в работах Т.А. Комаровой с соавторами [2, 3, 8]. Число модельных растений, взятых на временных пробных площадях, было недостаточным для того, чтобы сделать выводы о границах абсолютного возраста отдельных онтогенетических состояний, поэтому вблизи ППП 71-2012 и трех ВПП были собраны дополнительные образцы растений. В лабораторных условиях у каждого образца был подсчитан абсолютный возраст по числу годичных колец на уровне корневой шейки, а также число годичных колец на высоте 1,3 м (если растения достигали такой высоты).
Результаты исследования и их обсуждение
На ППП № 71-2012 растения P. koraiensis, P. ajanensis и A. nephrolepis представлены практически во всех онтогенетических состояниях (отсутствуют только сенильные растения P. ajanensis и A. nephrolepis) (рисунок). Онтогенетические спектры трех хвойных видов левосторонние, т.е. число растений быстро уменьшается от имматурных к виргинильным и генеративным (ось ординат представлена в логарифмическом масштабе). По спектрам всех видов заметно уменьшение числа растений до состояния v1, и затем их накопление до состояния v3. Далее у P. koraiensis и P. ajanensis число молодых генеративных растений вновь уменьшается и накапливаются они уже в среднегенеративном состоянии. У A. nephrolepis плавное снижение числа растений отмечается начиная с виргинильного состояния.
Такой характер онтогенетических спектров, вероятнее всего связан с жизненными стратегиями видов. Так, для P. koraiensis и P. ajanensis свойственно накопление растений в состоянии im1 (особи накапливаются под пологом древостоя, образуя группы), затем в v3 (поскольку при улучшении условий растения быстро проходят стадии v1 и v2) и в генеративном состоянии (g2, одна из самых длительных стадий), что связано с поэтапным выходом этих растений в полог древостоя. Для A. nephrolepis также характерно накопление имматурных растений, но за этим следует лишь однократное резкое увеличение прироста [9], а затем число растений постепенно уменьшается.
Спектры трех хвойных видов на временных пробных площадях неполночленные, но тем не менее они показывают, что группы подроста образованы растениями в разных онтогенетических состояниях (рисунок). При этом число особей P. koraiensis, A. nephrolepis и P. ajanensis существенно отличается на всех ВПП. Так, у A. nephrolepis на ВПП 1 максимальное число растений находится в состоянии im3 и не превышает 15 штук, на ВПП 2 – практически равномерное распределение особей во всех возрастных состояниях до v2 (около 10–12 шт. в каждом), а на ВПП 3 максимальное число растений находится в состоянии im2 (более 200 шт.). Растения P. ajanensis очень малочисленны на всех ВПП и лишь на ВПП 3 их число превышает десять штук. У P. koraiensis число имматурных особей первой подгруппы (im1) значительно больше, чем в других онтогенетических состояниях и колеблется от 75 (ВПП 2) до 140 штук (ВПП 3). Помимо имматурных растений в пятнах присутствуют и виргинильные: на ВПП 1 они есть у P. koraiensis и A. nephrolepis, на ВПП 2 и 3 – у всех видов.
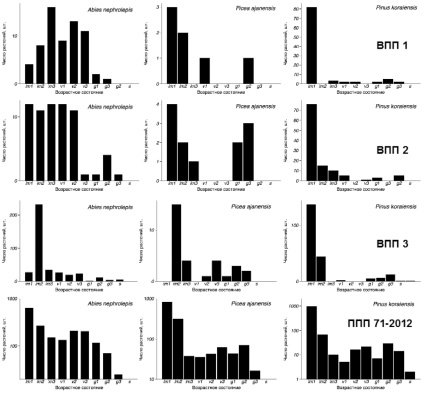
Возрастные спектры трех хвойных видов на постоянной и временных пробных площадях
Наличие особей в разном онтогенетическом стоянии на ППП 71-2012 (рисунок) может означать присутствие нескольких обособленных друг от друга групп (онтогенетических парцелл), внутри которых преобладают растения лишь в одном возрастном состоянии, как это представляется в gap-теории [1], но наличие особей разных онтогенетических состояний внутри групп возобновления (рисунок, ВПП 1, 2, 3) опровергает это предположение.
Подсчет годичных колец обнаруживает значительные вариации абсолютного возраста растений на временных пробных площадях. Так, у подроста P. koraiensis на ВПП 1 возраст колеблется от 0 (всходы) до 150 (v1) лет, на ВПП 2 – от 3 лет (juv) до 107 лет (im3), на ВПП 3 – от 4 лет (juv) до 119 лет (im3). У P. ajanensis на ВПП 1 – от 5 лет (im1) до 119 лет (v1), на ВПП 2 – от 24 лет (im1) до 120 лет (im3). У A. nephrolepis на ВПП 1 есть растения с возрастом от 7 лет (im1) до 120 лет (v1), на ВПП 2 – от 5 лет (juv) до 87 лет (v1). Большие различия в возрасте выявляются и внутри онтогенетического состояния и даже внутри отдельной подгруппы. На ВПП 1 у P. koraiensis есть растения в состоянии im1, возраст которых составляет от 16 до 48 лет. На ВПП 2 эти различия еще выше – от 15 до 54 лет. У P. ajanensis различия между имматурными растениями первой группы составили от 9 до 45 лет (ВПП 3). Для A. nephrolepis различия в одной группе возрастного состояния обычно не так значительны и составляют от 18 до 35 лет, но в то же время для нее характерно присутствие растений на ВПП во всех возрастных состояниях.
Сопоставление абсолютного возраста образцов растений, полученных на ВПП и в их окрестностях, показало, что особи одного онтогенетического состояния значительно отличаются по абсолютному возрасту, причем размах абсолютного возраста увеличивается от ювенильных растений к виргинильным (таблица). Например, уже у ювенильных растений P. koraiensis отличия в абсолютном возрасте достигают 17 лет. У имматурных растений отличия еще более значительны: до 48 лет у первой подгруппы, до 69 лет – у второй подгруппы и до 91 лет – у третьей подгруппы.
Абсолютный возраст растений в стадии подроста по возрастным состояниям
|
Возрастное состояние |
Pinus koraiensis |
Picea ajanensis |
Abies nephrolepis |
|||||||||
|
N |
Amin |
Amax |
∆A |
N |
Amin |
Amax |
∆A |
N |
Amin |
Amax |
∆A |
|
|
juv |
276 |
3 |
20 |
17 |
3 |
1 |
8 |
7 |
56 |
3 |
11 |
8 |
|
im1 |
385 |
6 |
54 |
48 |
208 |
2 |
49 |
47 |
670 |
7 |
39 |
32 |
|
im2 |
79 |
11 |
80 |
69 |
106 |
15 |
58 |
43 |
181 |
14 |
65 |
51 |
|
im3 |
72 |
16 |
107 |
91 |
40 |
20 |
110 |
90 |
73 |
28 |
100 |
72 |
|
v1 |
66 |
28 |
141 |
113 |
44 |
28 |
120 |
92 |
81 |
59 |
102 |
43 |
Условные обозначения: juv – ювенильное возрастное состояние, im1, im2, im3 – имматурные растения первой, второй, третьей группы, v1 – виргинильные растения первой группы; Amin,max – минимальный и максимальный абсолютный возраст в возрастном состоянии, лет; ∆А – вариация абсолютного возраста в возрастном состоянии, лет; N – число образцов, штук.
Исходя из представленных результатов можно сделать вывод, что группы подроста образованы растениями не только разного онтогенетического состояния, но и существенно разного абсолютного возраста. Принимая во внимание максимальный возраст виргинильных растений можно отметить, что размах абсолютного возраста в группах подроста в кедрово-широколиственных лесах достигает 100–140 лет.
Как было показано ранее [4, 9, 10], в случае улучшения условий (образование окна в пологе древостоя) имматурные растения ускоряют развитие и переходят в виргинильное состояние, достигая при возрасте 120–150 лет высоты 10–15 м. При отсутствии требуемого количества света они вновь способны приостанавливать развитие и некоторое время накапливаться, также формируя группы. Три рассматриваемых вида имеют различия в общей продолжительности жизни, продолжительности отдельных возрастных состояний [2, 3, 8] и способности переносить угнетение.
Если происходит образование окна, то разновозрастные имматурные растения хвойных видов одновременно увеличивают прирост и быстро занимают окно. Таким образом, зарастание окна обеспечивается накопленным ранее подростом. Если таких окон образуется несколько, то подрост из разных групп может сформировать новое поколение. При этом растения A. nephrolepis, с одной стороны, показывают большую теневыносливость и, с другой стороны, первыми увеличивают прирост. Потому они многочисленны, практически равномерно рассредоточены по площади и способны быстро заполнить образовавшееся окно. Реакция растений P. koraiensis на улучшение условий замедленная, поскольку особи нуждаются в существенно большем количестве света для развития. Поэтому их численность, начиная с состояния im2, относительно мала. Для растений этого вида важно достичь нижней части первого подъяруса, где они приостанавливают рост в высоту начинают накапливаться. Растения P. ajanensis, как и A. nephrolepis, сравнительно теневыносливы и потому могут формировать многочисленные группы в разных онтогенетических состояниях.
Заключение
Группы подроста, наблюдаемые под пологом древостоя в кедрово-широколиственных лесах, образованы растениями разного онтогенетического состояния и существенно различаются по абсолютному возрасту. Они представляют собой результат длительного накопления, обусловленного образованием небольших окон и прорывов в пологе, но не одновременного приживания растений. Растения трех хвойных видов P. koraiensis, P. ajanensis и A. nephrolepis на ранних стадиях онтогенеза в течение длительного периода времени способны переносить угнетение и дожидаться подходящих условий (образования окон) для дальнейшего развития. Поскольку группы подроста образуют мозаику, то они в значительной мере способны определять формирование мозаики и генеративных растений.
Рецензенты:
Баркалов В.Ю., д.б.н., заведующий отделом лесных и почвенных ресурсов, Биолого-почвенный институт ДВО РАН, г. Владивосток;
Денисов Н.И., д.б.н., главный научный сотрудник, Ботанический сад-институт ДВО РАН, г. Владивосток.
Работа поступила в редакцию 19.12.2013.
Библиографическая ссылка
Омелько А.М., Ухваткина О.Н., Жмеренецкий А.А., Гусев В.С. ВОЗОБНОВЛЕНИЕ ХВОЙНЫХ ВИДОВ В КЕДРОВО-ШИРОКОЛИСТВЕННЫХ ЛЕСАХ ЮГА РОССИЙСКОГО ДАЛЬНЕГО ВОСТОКА: ВОЗРАСТНАЯ СТРУКТУРА ПОДРОСТА // Фундаментальные исследования. – 2013. – № 11-5. – С. 943-947;URL: https://fundamental-research.ru/ru/article/view?id=33230 (дата обращения: 27.04.2024).