



Исследование электрической активности мозга млекопитающих при общей гипотермии имеет важное значение для выяснения механизмов работы мозга при низких температурах тела [7]. Электроэнцефалограмма (ЭЭГ) представляет собой результат суммации большого числа постсинаптических потенциалов большого числа нейронов. В мозге существует множество различных химических синапсов, отличающихся друг от друга природой нейротрансмиттера. Но выделение нейротрансмиттеров в химических синапсах в ответ на потенциал действия, поступающий в аксонную терминаль, инициируется потенциал-зависимыми кальциевыми каналами. В мозге существует много различных типов кальциевых каналов, отличающихся своими кинетическими свойствами, вольтамперной характеристикой и реакцией на изменения мембранного потенциала. В частности, в мозге существуют так называемые кальциевые каналы L-типа, которые реагируют на деполяризацию мембраны нейронов и участвуют в файринге нейронов. Эти каналы медленно инактивируются и, соответственно, пропускают большое количество ионов кальция [5]. Специфическим блокатором кальциевых каналов L-типа является нифедипин – производное дигидропиридина [10]. Нифедипин используется в качестве лекарства при гипертензии, и этот лечебный эффект обусловлен блокированием нифедипином кальциевых каналов L-типа. Ранее нами систематически исследовалась электрическая активность мозга крыс при гипотермии [1]. Нами было показано, что под тиопенталовым наркозом ЭЭГ крыс становится плоской при температуре тела 18–20 °С. Однако если перед охлаждением ввести животному мочевину в дозе 3 мМ на 100 г веса тела, то критическая температура тела, при которой ЭЭГ становится плоской, снижается до примерно 15 °С [2]. Механизм этого эффекта мочевины не известен. Некоторые структурные аналоги мочевины (например, ацетамид) также снижают критическую температуру [3]. Было предположено, что мочевина может отчасти блокировать кальциевые каналы, тем самым, предотвращая перегрузу нейронов кальцием при глубокой гипотермии.
В недавно опубликованной работе было показано, что нифедипин – блокатор кальциевых каналов L-типа – при введении в организм крысы увеличивает выделение глутамата в мозге крыс. Причём оказалось, что этот эффект не связан с влиянием нифедипина на кальциевые каналы L-типа. Глутамат является основным возбуждающим нейротрансмиттером в головном мозге [4]. Поэтому можно было ожидать, что введение нифедипина крысам перед общим охлаждением тела снизит критическую температуру для ЭЭГ.
Материалы и методы исследования
Животные. Опыты проведены на 12 крысах-самцах линии Вистар весом 180–200 г, содержащихся на обычном рационе в условиях вивария. Наркоз. Все опыты проведены на животных наркотизированных тиопенталом в дозе 40 мг на кг веса тела. Гипотермия. Снижение температуры тела осуществляли с помощью полиэтиленовых мешков, наполненных битым льдом. Наркотизированное животное помещали на мешки со льдом и периодически измеряли температуру тела в прямой кишке с помощью ртутного термометра. Время охлаждения составляло около 1,5 часа, а время согревания до нормотермии – около 1 часа. Электроды. ЭЭГ регистрировали с помощью нихромовых электродов диаметром 0,3 мм. Электроды имплантировали в соматосенсорную кору в область (примерные координаты (АР + 1, D2) и (AP + D3), у поверхности коры. Заземляющий электрод вживляли в носовую кость. Введение нифедипина. Нифедипин (фармацевтический препарат) вводили внутрибрюшинно в дозе 5 мг на 100 г веса тела.
Регистрация ЭЭГ. Регистрация ЭЭГ осуществлялась с помощью четырёхканального усилителя биопотенциалов, собранного в лаборатории биофизики биологического факультета ДГУ. Сигнал с выхода усилителя подавался на аналого-цифровой преобразователь (АЦП) ЛА-И24USB фирмы Руднев-Шиляев. С АЦП сигнал подавался на компьютер и записывался на жёстком диске. Частота оцифровки составляла 800 Гц. Длина записи 33 с (около 20 000 точек). Регистрацию производили по мере снижения температуры тела через каждые два градуса. После достижения критической температуры тела, когда ЭЭГ становилась изоэлектрической, охлаждение прекращали и начинали согревание. Для этого под тело животного подкладывали грелку с тёплой водой. ЭЭГ регистрировали по мере повышения температуры тела через каждые два градуса. Анализ ЭЭГ. После конвертирования данных их сохраняли в формате пакета STATISTIKA. Далее использовались программы опции «анализ временных рядов». Строили графики самой ЭЭГ (2,5-секундные (2000 точек) отрезки ЭЭГ и её спектр мощности. Спектральная плотность выражалась в процентах от общей мощности в диапазоне частот от 0 до 15 Гц, принятой за 100 %.
Результаты исследования и их обсуждение
На рис. 1 приведены электроэнцефалограммы крыс c введением нифедипина в дозе 5 мг/100 г веса тела. По мере снижения температуры тела амплитуды колебаний на ЭЭГ уменьшаются. Спектр мощности ЭЭГ закономерно изменяется (рис. 2). При температуре тела 32 °С появляется полоса в области 6–7 Гц. При дальнейшем снижении температуры тела она смещается в область низких частот. При температуре тела 15 °С ЭЭГ становится практически плоской. Таким образом, введение нифедипина существенно снижает температуру исчезновения электрической активности. При согревании животного после глубокой гипотермии ЭЭГ восстанавливается при температуре тела 21–23 °С. Однако колебания на ЭЭГ нерегулярные, амплитуда низкая.
При температуре тела 36 °С колебания регулярные, амплитуды колебаний примерно такие же, как и до начала охлаждения. Спектр мощности ЭЭГ имеет примерно такой же вид, как и до начала охлаждения. Хотя и видно, что эти спектры отличаются друг от друга. Например, мощность колебаний в области частот от 0 до 3 Гц в начале эксперимента заметно выше, чем после согревания, хотя температура тела в обоих случаях одинакова.
Диапазон частот, в котором появляется доминирующая полоса в спектре мощности, соответствует тета-ритму. Это значит, что при поверхностной (mild) гипотермии в коре головного мозга доминирует тета-ритм. Одной из причин такой картины может быть подавление активности большого числа корковых нейронов. Известно, что несинхронизированная активность большого числа нейронов приводит к высокочастотной низкоамплитудной картине ЭЭГ. Соответственно, спектр мощности будет содержать широкий набор частот. Известный эффект возникновения альфа-ритма в зрительной коре при закрывании глаз объясняется именно синхронизацией активности нейронов. Но синхронизация невозможна при активности большого числа нейронов. Поскольку нейроны образуют сеть взаимосвязанных генераторов потенциалов действия. Когда сенсорная информация блокируется, импульсация в соответствующие проекции коры перестаёт поступать и большое число нейронов перестаёт генерировать потенциалы действия. Тогда становится возможной синхронизация работы «опорных нейронов», генераторов ритма. Тогда высокочастотные колебания на ЭЭГ исчезают, а ритм пейсмейкера становится доминирующим.
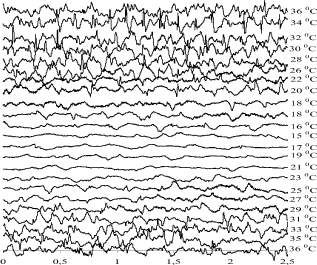
Рис. 1. ЭЭГ крысы при охлаждении и последующем согревании при введении нифедипина. Масштаб по вертикали 400 мкВ, по горизонтали – 1 с
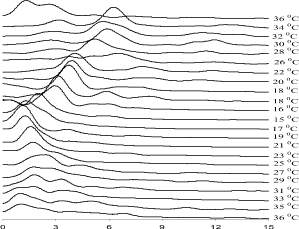
Рис. 2. Спектральные плотности ЭЭГ крысы при охлаждении и последующем согревании при введении нифедипина. По оси абсцисс – частота в Гц, по оси ординат спектральная плотность в процентах от общей плотности в диапазоне от 0 до 15 Гц
Известно, что амплитуда постсинаптического ответа существенно зависит от количества медиатора, выделяющегося в щель в ответ на приходящий в терминаль потенциал действия [9]. В свою очередь, количество выделяющегося в синаптическую щель медиатора определяется вероятностью наступления событий, ведущих к образованию поры, через которую медиатор поступает из синаптического пузырька в синаптическую щель Известно, что ионы кальция играют важную роль в процессе выделения медиатора в синаптическую щель. В ответ на приходящий в терминаль потенциал действия ионы кальция через потенциалзависимые кальцивые каналы поступают из экстраклеточного пространства в терминаль, где связываются с белками специализированного комплекса, осуществляющего процесс образования поры в пресинаптической мембране и выделения медиатора в синаптическую щель. Исследование зависимости количества выделившегося в синаптическую щель медиатора от концентрации экстраклеточного кальция указывает на то, что по крайней мере четыре иона кальция должны связаться с медиаторвыделяющим белковым комплексом для того, чтобы запустить процесс сплавления мембраны синаптического пузырька с пресинаптической плазматической мембраной и выделения медиатора в синаптическую щель [6]. Это означает, что количество выделяющегося в ответ на ПД медиатора существенно зависит от концентрации кальция в месте локализации медиаторвыделяющего комплекса. Например, если вероятность выделения медиатора зависит в четвертой степени от концентрации кальция, то изменение эффективной концентрации кальция всего лишь на 20 % изменит количество выделяемого медиатора в два раза. Поэтому реакции, влияющие на эффективную концентрацию кальция, будут существенно влиять на количество выделяющегося медиатора и на амплитуду постсинаптических потенциалов. Эффективная концентрация кальция в месте локализации медиатор выделяющего комплекса зависит:
- от концентрации экстраклеточного кальция;
- проводимости и времени открытого состояния кальциевого канала;
- емкости кальциевого буфера в окрестности пресинаптических пузырьков, адсорбированных на пресинаптической мембране.
Все эти величины зависят от температуры, но надо полагать, что наибольшей чувствительностью к температуре обладают константы скорости открытия и закрытия кальциевого канала. В работе [8] показано, что Q10 ~ 2,5, если предположить, что эта закономерность имеет общий характер, то снижение температуры должно увеличивать количество кальция проходящего через канал в ответ на ПД, а следовательно, и амплитуду постсинаптического потенциала. Увеличение амплитуды ПСП увеличивает вероятность генерации ПД в районе аксонного холмика, а следовательно, и генерации последующего ПСП. Предельная частота, которую может пропустить синапс, зависит от концентрации экстраклеточного кальция: чем меньше эта концентрация, тем выше предельная частота [9]. Можно предположить, что введение нифедипина увеличивает предельные частоты для синаптических контактов и тем самым увеличивает среднюю частоту ЭЭГ.
Возможно, введение нифедипина уменьшает эффективную концентрацию кальция в экстраклеточном пространстве или уменьшает вход кальция в пресинаптическую терминаль в ответ на потенциал действия. Это, в свою очередь, уменьшит количество выделяющегося медиатора в ответ на потенциал действия. Таким образом, каждый потенциал действия будет расходовать меньше «ресурсов» по выражению Цодыка и Маркрама.
Рецензенты:
Омаров К.З., д.б.н., профессор, зав. лабораторией экологии животных, ФГБУН ПИБР ДНЦ РАН, г. Махачкала;
Габибов М.М., д.б.н., профессор, зав. кафедрой анатомии, физиологии, гистологии, ГОУ ВПО ДГУ, г. Махачкала.
Работа поступила в редакцию 28.05.2014.
Библиографическая ссылка
Абдурахманов Р.Г., Пиняскина Е.В., Пиняскина Е.В., Гитиномагомедовa М.М. ВЛИЯНИЕ НИФЕДИПИНА НА ЭЛЕКТРИЧЕСКУЮ АКТИВНОСТЬ МОЗГА КРЫС ПРИ ГИПОТЕРМИИ // Фундаментальные исследования. – 2014. – № 8-3. – С. 620-623;URL: https://fundamental-research.ru/ru/article/view?id=34604 (дата обращения: 26.04.2024).