



Различные формы численных и структурных аномалий хромосом (геномных вариаций), были уже неоднократно ассоциированы с нарушениями психики [1–7, 22–27, 33, 39, 46–48]. В случаях таких социально значимых заболеваний, как идиопатические формы умственной отсталости и аутизма, а также при шизофрении, геномные вариации обнаруживаются в 30–50 %, 5–40 % и 1–10 % случаев соответственно [3, 4, 39, 47, 51]. Численные аномалии хромосом, или анеуплоидия, как правило, являются маркерами патологических состояний и довольно часто несовместимы с жизнью [1, 43–45]. Анеуплоидия, которой предшествует каскад аномальных митотических делений, также обнаруживается практически при всех онкологических заболеваниях [1, 9, 23]. Более того, нарушение функционирования аппарата митотического деления, вероятно, ассоциировано со старением [9, 53]. Анеуплоидия аутосом является одной из наиболее частых форм численных хромосомных аномалий, выявляемых в материале спонтанных абортов и у мертворожденных детей. Частота хромосомных аномалий, обнаруживаемых в спонтанных абортусах, была оценена как 50–70 % [44, 45]. Регулярные формы таких хромосомных аберраций являются результатом нерасхождения хромосом в мейозе, что ведет к появлению гамет с аномальным хромосомным набором. Мозаичные формы аномалий хромосом, напротив, возникают вследствие постмейотических ошибок [24]. Мейотический процесс чувствителен к влиянию эндогенных и экзогенных факторов. Возраст родителей, влияние алкоголя, табака, других мутагенных субстанций и подверженность радиации являются одними из наиболее широко известных причин, ассоциированных с нерасхождением хромосом в митозе. Исследования, проведенные на больших когортах и посвященные изучению ассоциации частоты синдрома Дауна и возраста матери (в случаях, где причина заболевания – нерасхождение хромосом 21 в материнском мейозе I или II), выявили корреляцию между этими двумя факторами [12–17, 34]. В исследовании 74 ядерных семей больных с синдромом Дауна было выявлено то, что нерасхождение в материнском мейозе было характерно для 58 семей (86 %), в отцовском – для 7 семей (10 %). Из доступных для анализа случаев материнское нерасхождение в I мейозе наблюдалось в 43 (64 %) случаях, во II – в 15 (22 %), нерасхождение в отцовском I мейозе – 2 (3 %), II – 5 (7 %) (рис. 1) [42]. Антенатальную гибель плода, как правило, связывают с трисомией хромосом 13, 18 и 21, тогда как спонтанные аборты в большей степени ассоциированы с трисомиями хромосом 2, 4, 7–9, 13–16, 18, 20–22 [1, 39, 41]. Показано, что большинство форм аутосомной анеуплоидии не совместимо с нормальным внутриутробным развитием [1, 31, 32, 43].
Цитогенетические исследования новорожденных и детей с пороками развития демонстрируют преобладание в данной группе трисомии хромосомы 21 (синдром Дауна), намного реже – трисомии хромосом 18 и 13 (синдромы Эдвардса и Патау, соответственно), в отдельных случаях – мозаичные формы численных аномалий других аутосом [1]. Трисомия хромосомы 21, являющаяся причиной более чем 95 % случаев синдрома Дауна, остается значимой генетической причиной умственной отсталости и обнаруживается примерно у 1 из 600–800 живорожденных детей. Рожденные с синдромом Дауна представляют небольшое число из всех плодов с трисомией хромосомы 21, так как более 80 % с этим синдромом погибают внутриутробно, вызывая 1–2 % спонтанных абортов [10, 40]. Моносомия хромосомы 21 была обнаружена у живорожденного ребенка лишь в нескольких случаях, однако врожденные пороки развития, несовместимые с жизнью, влекли за собой гибель в течение первого дня после рождения [32, 43]. Следует отметить, что, несмотря на очевидно негативный эффект анеуплоидии в клетках крови, имеются предположения о том, что повышенные уровни анеуплоидии характерны для нормально развивающегося мозга плода. Изучение генетических аномалий головного мозга (не рассматривая онкологические заболевания) началось сравнительно недавно, в 2000 году [7, 23, 55]. Головной мозг взрослого человека в основном состоит из нейронов, которые являются постмитотическими, неделящимися клетками. Исследования хромосомного набора в клетках головного мозга были невозможны ввиду недоступности методов визуализации, позволяющих исследовать интерфазные ядра.
До появления интерфазного анализа считалось, что в головном мозге здорового человека, содержащем примерно 1012 нейронов, хромосомный набор должен быть нормален и идентичен для всех клеток, как и в остальных тканях [7, 22, 27, 49–55]. Однако после проведения исследований головного мозга мышей были получены первые данные о различных формах соматических, геномных и хромосомных вариациях нейронов головного мозга [8, 23, 55]. С помощью флюоресцентной гибридизации in situ (FISH) и спектрального кариотипирования нейробластов коры головного мозга в развивающемся и взрослом мозге мышей было выявлено то, что более 30 % нейробластов являются анеуплоидными. Визуализация метафазных хромосом также определила, что для многих нейронов коры головного мозга был характерен аномальный кариотип. Соответственно в головном мозге мышей были выявлены геномные вариации на хромосомном уровне в развивающихся и сформировавшихся нейронах [37].
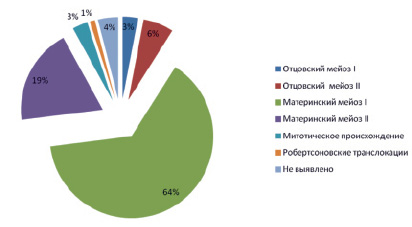
Рис. 1. Частота хромосомных нарушений при синдроме Дауна [12, 13, 42]
Сложно представить, что геном всех клеток человека одинаков. Организм взрослого человека состоит как минимум из 1014 клеток 210 различных типов. Все они берут начало от одной диплоидной клетки (зиготы), которая делится путем митоза примерно 1016 раз, приводя к образованию 45 поколений клеток. Геномные вариации в соматических клетках присутствуют постоянно как результат более 100 повреждений ДНК в день. Большая часть таких повреждений нейтрализуется системой репарации ДНК, однако в то же время эти процессы представляют собой источник экзогенного изменения генома и могут быть причиной различных патогенных процессов [27]. Несмотря на это, известно, что хромосомная нестабильность, ограниченная определенной тканью, может не приводить к нарушению ее функционирования [5, 15, 25, 29]. Было показано, что феномен хромосомного мозаицизма, ограниченного плацентой, идентифицируется приблизительно в 1–2 % нормальных беременностей [30], а в эмбриональных тканях яичника отмечаются повышенные уровни анеуплоидии с вовлечением хромосомы 21 [16]. Тем не менее во многих работах были отмечены ассоциации между наследственными заболеваниями и мозаичными вариациями числа копий (CNV) [26, 33]. Мозаичные формы численных хромосомных аномалий наблюдаются при хромосомных синдромах в 3–18 % случаев [1, 27, 39]. В литературе также описано большое количество случаев мозаичных численных и структурных хромосомных аномалий, при которых наблюдались умственная отсталость или другие нарушения психики, врожденные пороки развития, аутоиммунные болезни, а в некоторых случаях отсутствие клинических проявлений [1–7, 24–27, 39, 47]. Принимая во внимание и то, что возникновение мозаичных клонов, как предполагается, происходит в период внутриутробного развития, а также тот факт, что среди образцов, используемых для пренатальной диагностики, частота мозаицизма значительно выше по сравнению с постнатальным периодом, необходимо рассмотреть хромосомный мозаицизм в головном мозге в ходе онтогенеза. Первая попытка оценить число хромосом в мозге человека была предпринята группой ученых в 1991 году. Хотя ученые исследовали генетические изменения в опухолях мозга, при этом одновременно был выявлен высокий уровень трисомии у людей из контрольной группы (средний уровень – 2 % на отдельную хромосому) (для более детального описания этих работ см. [23]). Дальнейшие работы, проведенные на интерфазных ядрах головного мозга, позволили сравнить уровни анеуплоидии у плода и взрослого индивидуума, оценить различия по уровням для отдельных хромосом у здоровых людей и у пациентов с нервно-психическими, онкологическими и другими заболеваниями, а также в некоторых случаях определить пороговые значения в норме и патологии [18–23, 37, 46, 48–55]. Так, при исследовании анеуплоидии в клетках головного мозга плода было обнаружено то, что анеуплоидия выявляется почти на всех этапах внутриутробного развития и достигает наивысшего уровня в первом триместре беременности [49, 50]. В одной из современных работ по данной теме описано исследование 12 образцов ткани мозга плода, полученных после прерывания беременности, не связанной с генетической причиной. После исследования хромосом 1, 18, 13/21, X и Y было выявлено, что частота ядер с аномальным хромосомным набором составляет от 0,2 % до 4,4 % в некультивированных клетках мозга, уровень анеуплоидии для различных хромосом варьировался от 0,4 % до 11 % [18, 21, 49]. Однако в мозге взрослого индивидуума средний уровень анеуплоидии на хромосому находится в диапазоне от 0,1 до 0,7 %, с общей частотой анеуплоидии по всем хромосомам близкой к 10 %, что в три раза меньше, чем в эмбриональном мозге [21]. Вероятно, такое снижение количества анеуплоидных клеток происходит за счет запрограммированной смерти аномальных клеток с хромосомными и геномными нарушениями, вследствие чего и наблюдается уменьшение уровня нестабильности (анеуплоидии) в три раза в клетках головного мозга, согласно исследованиям постнатального мозга [20, 28, 50]. В мозге здорового взрослого индивидуума, как уже было отмечено, выявлен определенный уровень анеуплоидии по отдельным хромосомам [27, 53–55]. Исходя из того, что повышенные уровни анеуплоидии хромосом в головном мозге, обнаруживаемые пренатально, не выявляются у индивидуумов во взрослом возрасте, можно предположить, что для нормального развития центральной нервной системы анеуплоидия должна быть элиминирована [50]. Вероятно, элиминация анеуплоидных клеток в головном мозге замедляется с возрастом, что может быть ассоциировано со старением и малигнизацией. Это предположение поддерживается результатами последних исследований старения и онкологических заболеваний [23, 25, 38, 52, 53]. Предполагается, что наличие высокого уровня анеуплоидии в головном мозге характерно для онкологической патологии мозга и нервно-психических заболеваний (рис. 2).
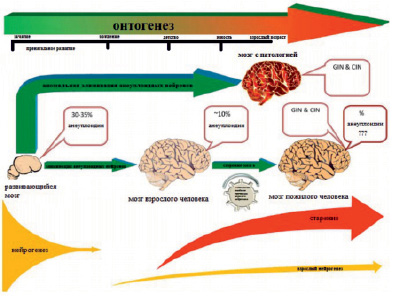
Рис 2. Анеуплоидный нейрон головного мозга может быть вовлечен в сети нейронального взаимодействия. Воздействуя на элементы нейрональных сетей через синаптическую активность, такой нейрон может продуцировать нарушения в клетках головного мозга либо быть связан с механизмом старения. Если уровень анеуплоидии относительно невелик, интеграция анеуплоидного нейрона в нейрональные сети может быть связана с механизмом нейронального разнообразия [53]
В данном контексте особый интерес представляет изучение клеток головного мозга при нервно-психических заболеваниях и выявление отличий уровня анеуплоидии от контрольной когорты. Группой авторов было проведено исследование образцов префронтальной коры (область 10, серое вещество), полученных от шести пациентов с шизофренией и двух пациентов из контрольной группы [55]. На момент смерти пациентам с шизофренией было от 22 до 77 лет, пациентам из контрольной группы – от 28 до 76 лет, время с момента смерти и до получения образца – 2–6,5 часа и 3–7 часов соответственно. Причины смерти в обоих группах – инфракт, пневмония, эмболия легких. Было проанализировано 200 нейронов каждого образца мозга (по 200 ядер на каждую пробу ДНК). Большинство нейронов содержало две копии хромосом 1, 7, 8, 13, 16, 18, 21, 22, X (женщины) и XY – мужчины, однако у двух индивидуумов с шизофренией были обнаружены повышенные уровни анеуплоидии с вовлечением хромосом Х и 18. У одного пациента было обнаружено 4 % клеток с трисомией хромосомы Х и 2,5 % – с трисомией хромосомы 18, у другого – 3 % клеток с трисомией Х и 0,5 % – с трисомией 18; у остальных индивидуумов, как с шизофренией, так и из контрольной группы, трисомии по исследованным хромосомам обнаружено не было [55]. Исходя из этого, можно предположить, что при нервно-психических заболеваниях, судя по всему, наблюдается увеличение уровней анеуплоидии по отдельным хромосомам, поэтому изучение геномной нестабильности в клетках головного мозга и лимфоцитах периферической крови у пациентов с нервно-психическими заболеваниями представляет собой непосредственный интерес для поиска вероятных маркеров этих заболеваний.
В настоящее время связь повышенного уровня анеуплоидии определенной хромосомы в головном мозге с конкретным заболеванием пока не определена, хотя очевидно, что исследования только культуры лимфоцитов периферической крови пациентов с нервно-психическими заболеваниями могут быть недостаточны для шизофрении и болезни Альцгеймера, поскольку при этих болезнях анеуплоидия может быть специфичной для клеток головного мозга. По-видимому, повышенные уровни анеуплоидии разных хромосом связаны с функциональными нарушениями мозга, однако для подтверждения этого предположения необходимо рассматривать каждую хромосому в отдельности, как в контрольной группе, так и у пациентов с нарушениями психики для определения их вклада в патогенез заболевания. В качестве одной из самых значимых для нормального функционирования мозга можно считать хромосому 21. Как известно, трисомия хромосомы 21 является трисомией аутосомы, при которой плод может не только выжить после рождения, но и жить продолжительной жизнью по сравнению с другими самыми распространенными трисомиями (трисомии хромосом 18 и 13). Анализируя анеуплоидию хромосомы 21 в клетках головного мозга (постмортальные образцы) у здоровых индивидуумов и пациентов с нервно-психическими заболеваниями (таблица), в первую очередь, стоит обратить внимание на уровни анеуплоидии, полученные для контрольных групп в различных исследованиях [4–7, 19–29, 46, 47, 51–55]. В одной из работ авторы проанализировали более 5000 интерфазных ядер двух различных соматических тканей – ворсины хориона, постмортальные клетки мозга плода и взрослых индивидуумов. Было отмечено, что интерфазные трисомные ядра составляют от 0,3 до 0,7 %, при этом в ворсинах хориона – от 0,4 % до 0,7 %, головном мозге плода – 0,3 %, в постмортальных образцах мозга детей – 0,3 %, взрослых – 0,42 % [50]. Исследования анеуплоидии хромосомы 21 в клетках головного мозга показали, что средний уровень анеуплоидии этой хромосомы варьируется в пределах от 0,2 до 4 %. Для сравнения, уровень анеуплоидии по хромосоме 21 в интерфазных культивируемых лимфоцитах был определен как ~ 0,6 %. Хотя результаты этой работы не позволяют оценить уровень анеуплоидии в головном мозге, так как была проанализирована только одна хромосома, однако они позволяют предположить, что по определенным хромосомам средний уровень анеуплоидии может быть выше в одних исследованиях по сравнению с другими [16, 21, 23, 37]. Помимо этого, в ряде работ исследовались уровни анеуплоидии для хромосом 1, 7, 14 и 21 с помощью интерфазного многоцветового хромосомоспецифичного окрашивания (ICS-MCB). Средний уровень анеуплоидии для этих хромосом был определен в 1,4–2,6 % (с пороговыми значениями от 2,0 до 3,8 %). Из описанных исследований очевидно, что данные о среднем уровне анеуплоидии для хромосомы 21 немного расходятся, но в ранних исследованиях все результаты по анеуплоидии выше 4 % свидетельствовали о повышении уровня анеуплоидии и, вероятно, об аномальных процессах в головном мозге [5–7, 18–22, 50, 51, 54]. В одной из работ [21] были исследованы клетки головного мозга пациентов с болезнью Альцгеймера и обнаружено увеличение уровня анеуплоидии, специфичной для хромосомы 21, до 11 %. Ассоциация болезни Альцгеймера со специфичной для клеток головного мозга анеуплоидией хромосомы 21 отмечалась и в нескольких других работах [11, 35, 36]. Значительно повышенный уровень анеуплоидии хромосомы 21 в коре головного мозга позволил предположить, что данный феномен связан с нейродегенерацией при болезни Альцгеймера [21]. Хотя причины повышенного уровня анеуплоидии хромосомы 21 в мозге больных болезнью Альцгеймера остаются в значительной степени неясными, последние исследования показывают, что анеуплоидные нейроны, скорее всего, являются более предрасположенными к селективной клеточной смерти при болезни Альцгеймера [38]. Стоит также отметить, что анализ фибробластов кожи методом FISH показал наличие статистически значимого числа клеток с дополнительной хромосомой 21. При анализе клеток мозжечка пациентов с болезнью Альцгеймера с помощью FISH/QFISH и ICS-MCB ученые обнаружили значительное увеличение (в 10 раз) уровня специфичной анеуплоидии хромосомы 21 в клетках мозжечка пациентов, который был вовлечен в прогрессирующую дегенерацию (6–15 % по сравнению с 0,8–1,8 % в контрольной группе) [21, 28]. Иными словами, при болезни Альцгеймера выявляются соматические вариации генома в виде низкопроцентного хромосомного мозаицизма по анеуплоидии хромосомы 21 [28, 52]. Для сравнения результатов анализа клеток головного мозга пациентов с болезнью Альцгеймера с другими болезнями мозга, а также с контрольной группой были проанализированы препараты клеток головного мозга с атаксией-телеангиэктазией и пациентов без психических заболеваний. Были исследованы поле Бродмана 10 для пациентов с болезнью Альцгеймера и поле Бродмана 24 для индивидуумов с атаксией-телеангиэктазией.
Анеуплоидия хромосомы 21 в клетках головного мозга при нервно-психических заболеваниях
|
Заболевание |
Форма анеуплоидии |
Ссылки |
|
Болезнь Альцгеймера |
Мозаичная анеуплоидия хромосомы 21 (хромосомоспецифическая нестабильность), 6–15 % |
[22, 26, 28, 52] |
|
Атаксия-телеангиэктазия |
Мозаичная анеуплоидия и хромосомоспецифическая нестабильность, 2,3–3,1 % (для хромосом 1, 7, 14, 21 и X) |
[19, 21] |
|
Шизофрения |
Не выявлена |
[51, 55] |
Анализ хромосом 1, 7, 14 и 21 в 7000 ядрах 7 образцов головного мозга с помощью ICS-MCB выявил средний уровень анеуплоидии у индивидуумов без психических заболеваний, который примерно равен 1,4–2,6 % (с пороговыми значениями 2,0 и 3,8 %). Эти данные демонстрируют то, что процент стохастической анеуплоидии на пару хромосом (аномальные значения) в контрольных образцах стоит расценивать как увеличенный, т.е соответствующий мозаичной клеточной линии. Средний уровень анеуплоидии после исследования на пяти хромосомах (хромосомы 1, 11, 17, 18, X и Y) был определен как 0,5 %, с общим числом анеуплоидных клеток, приближающимся к 10 %. Исследование примерно 100000 интерфазных ядер пациентов с болезнью Альцгеймера, проведенное с помощью ICS-MCB, показало отсутствие значимого увеличения уровней анеуплоидии для хромосом 1, 7, 14 и X по сравнению с контрольной группой (p > 0,05; 2,5 и 3,6 % для пациентов c болезнью Альцгеймера по сравнению с 0,8–2,0 % в контрольной группе). Однако уровень анеуплоидии по хромосоме 21 в мозге пациентов с болезнью Альцгеймера составил 6–15 % по сравнению с 0,8–1,8 % в контрольной группе. Различие между этими результатами являлось статистически значимым (p < 0,001). В группе с атаксией-телеангиэктазией тоже был выявлен повышенный уровень анеуплоидии по хромосоме 21, однако он был сравним с увеличением уровней по другим хромосомам (уровень анеуплоидии для хромосом 1, 7, 14, 21 и X – 2,3 % – 3,1 %) [19, 21].
Вероятно, анеуплоидия хромосомы 21 в клетках головного мозга при болезни Альцгеймера является неотъемлемой составляющей патогенного каскада нейродегенерации [11, 36]. Подтверждением этому предположению может служить тот факт, что, так как хромосома 21 также кодирует ген белка-предшественника амилоида, все индивидуумы с синдромом Дауна демонстрируют признаки болезни Альцгеймера, и у большинства развивается деменция к 30–40 годам [14, 34–36, 38, 40]. Связь между синдромом Дауна и болезнью Альцгеймера была впервые описана два десятилетия назад [36]. У пациентов с синдромом Дауна в возрасте около 30 лет были выявлены изменения в головном мозге, которые гистологически неотличимы от обнаруживаемых у пациентов с болезнью Альцгеймера [11, 35, 36]. Известно, что ген АРР, отвечающий за продукцию белка-предшественника бета-амилоида, находится на хромосоме 21 в участке q21, поэтому предполагается, что трисомия хромосомы 21 может привести к увеличению синтеза этого белка и соответственно к увеличению продукции бета-амилоидного пептида, избыточное количество которого наблюдается при болезни Альцгеймера. Тот факт, что у пациентов с синдромом Дауна, достигших возраста 30 или 40 лет, неврологические нарушения в точности повторяют клиническую картину при болезни Альцгеймера, обеспечивает вклад в понимание патогенеза болезни Альцгеймера, акцентируя внимание на нерасхождении хромосомы 21, где находится ген APP, и на последствиях его измененной экспрессии. Биохимические и генетические исследования показывают, что как в спорадических, так и в семейных случаях болезни Альцгеймера, у пациентов нарушен тот или иной этап клеточного цикла [35]. Дополнительная копия хромосомы 21, которая при регулярной трисомии в случае синдрома Дауна приводит к нейродегенерации и деменции, может быть причиной как генетической, так и спорадической формы болезни Альцгеймера, в зависимости от того, были ли хромосомная нестабильность и мозаичная анеуплоидия вызваны генетической мутацией или неблагоприятным воздействием окружающей среды. Кроме того, предполагается, что нарушение функции микротрубочек вносит вклад в процесс аномального митоза и геномной нестабильности [11]. Синдром Дауна как модель нейродегенерации позволил исследователям сделать некоторые предположения касательно гипотез болезни Альцгеймера.
Таким образом, вероятно, определенное число клеток в тканях пациентов с болезнью Альцгеймера должно быть анеуплоидным, в том числе должна присутствовать трисомия хромосомы 21 в клетках соматических тканей. Изменение стабильности генома и появление мозаичной трисомии по хромосоме 21 будет способствовать началу деменции и нейродегенерации, но более медленными темпами, чем при синдроме Дауна. Мутации, приводящие к семейной форме болезни Альцгеймера, скорее всего происходят в генах, кодирующих белки, непосредственно или косвенно участвующие в клеточном цикле и сегрегации хромосом. К анеуплоидии при болезни Альцгеймера, в том числе мозаичной трисомии хромосомы 21, могут приводить изменения в микротрубочках, аппарате митотического деления и белках, связанных с митозом.
Заключение
Большое количество связей между нейронами головного мозга дает возможность предположить, что присутствие даже малого числа клеток с аномальным геномом может оказывать неблагоприятное влияние на весь организм и особенно на функционирование центральной нервной системы. Хотя роль анеуплоидии клеток головного мозга в формировании индивидуальных различий у здоровых людей еще не определена, но, тем не менее, предполагается, что повышенный уровень анеуплоидии различных хромосом характерен для определенных заболеваний головного мозга [19–23, 51–55]. Исследования анеуплоидии хромосомы 21 в клетках головного мозга у пациентов с болезнями мозга и в контрольных группах позволили установить пороговый уровень анеуплоидии для здоровых индивидуумов (не более 4 %), а также выявить увеличение уровня анеуплоидии (до 6–15 %) по данной хромосоме в мозге пациентов с болезнью Альцгеймера. Полученные результаты являются хромосомоспецифичными именно для данного заболевания и не наблюдаются при других нервно-психических заболеваниях, таких как атаксия-телеангиэктазия и шизофрения. В качестве основных причин приводящих, вероятно, к изменению клеточного цикла нейронов, называют возвращение постмитотического нейрона в цикл клеточного деления и невозможность полного прохождения (завершения) клеточного цикла. Пока не определено, является ли анеуплоидия ответом на нейродегенерацию, или же присутствует в мозге еще до ее начала. Несмотря на то, что описанные исследования позволяют сделать предположение о патогенном влиянии повышенного уровня анеуплоидии хромосомы 21 на функционирование головного мозга, их число недостаточно для более глубокого понимания роли анеуплоидии данной хромосомы. Мозаичная анеуплоидия в головном мозге может рассматриваться в качестве предполагаемого механизма генетического разнообразия или вариабельности генома, как в здоровом мозге, так и в мозге пациентов с нервно-психическими заболеваниями [22]. Для более полного представления о связи анеуплоидии клеток головного мозга с болезнями мозга необходимы более подробные исследования межхромосомных вариаций как нейрональных, так и глиальных клеток. Дополнительные исследования хромосомы 21, проведенные на клетках головного мозга пациентов с болезнью Альцгеймера, синдромом Дауна и в контрольных группах, возможно, будут способствовать более полному пониманию патогенеза заболеваний головного мозга.
Исследования анеуплоидии в клетках головного мозга в лабораториях авторов выполняются за счёт гранта Российского Научного Фонда (проект № 14-35-00060).
Библиографическая ссылка
Юров И.Ю., Ворсанова С.Г., Зеленова М.А., Юров Ю.Б. АНЕУПЛОИДИЯ ХРОМОСОМЫ 21 ПРИ ЗАБОЛЕВАНИЯХ ГОЛОВНОГО МОЗГА // Фундаментальные исследования. – 2015. – № 7-1. – С. 61-70;URL: https://fundamental-research.ru/ru/article/view?id=38726 (дата обращения: 26.04.2024).