



В процессе укоренения черенков (эксплантов) древесных растений большое значение наряду с другими факторами имеют: возрастное состояние, темпы развития маточного (исходного) растения и физиолого-биохимические особенности побегов, с которых берут черенок (эксплант) [4].
Известно, что органогенез в культуре тканей и клеток управляется с помощью определенных сочетаний фитогормонов (в том числе ауксинов и фенольных соединений) [1,7-8]. Рострегулирующие вещества играют важную роль в становлении организованной многоклеточной структуры растений, возникновении и развитии органов, они обусловливают дифференциальное выражение признака в фенотипе [2, 4]. Под их контролем находятся физиологические процессы и генетические программы, отвечающие за деление, растяжение клеток, дифференциацию побегов, корней и т.д.
Существует мнение, что изучение генетического контроля дифференцировки растений связано с идентификацией и характеристикой генов, обуславливающих фитогормональный статус [5]. Важным компонентом упорядочения, согласования и объединения структур и функций в целостном организме растений является индолилуксусная кислота (ИУК). Индолилуксусная кислота участвует в индукции и регуляции деления, растяжения, дифференциации, поляризации и питания клеток, в формировании базально-апикального рисунка в раннем эмбриогенезе, формировании корневых волосков, росте гипокотиля, тропизме, формировании листьев [1, 4, 6]. По мнению многих исследователей в процесс поддержания ауксинового гомеостаза вовлечено множество генов, что указывает на значимость этого гормона в развитии растений [5].
Большое значение в жизни растений имеют фенольные соединения, принимающие участие в окислительном метаболизме растений (в качестве доноров и акцепторов водорода), в биосинтезе индолилуксусной кислоты и в окисляющих её системах [9]. Некоторые из полифенолов и родственных им соединений – ингибиторы прорастания семян и роста растений.
Целью исследования являлось изучение физиолого-биохимических особенностей исходных для культивирования in vitro побегов деревьев дуба колонновидного (пирамидального) и черешчатого. В задачу исследований входило определение свободной (физиологически активной) фракции эндогенных регуляторов роста индольной (ауксины) и фенольной (ингибиторы роста) природы.
Материалы и методы исследования
Объектом исследований являлись клоны дуба черешчатого (Quercus robur L.) (прививка 1978 г.) и колонновидного (Quercus robur var. pyramidales Gmelin) (прививка 1965 г.). Были исследованы три дерева (раметы): одно дерева дуба черешчатого (поздняя форма, дерево № 56, произрастающее в Семилукском питомнике (Воронежская область) на лесосеменной плантации, созданной Ю.П. Ефимовым в 1976 г.) и два дерева дуба колонновидного (поздняя форма, прививка проведена В.К. Ширниным), произрастающих в г. Воронеже (дерево № 1 – Кольцовский сквер, дерево № 2 – на ул. Карла Маркса).
Рострегулирующие вещества определяли в побегах текущего года. Для анализа использовали первый и второй приросты. Пробы отобраны в периоды: интенсивного роста в длину (24.05.2011 и 16.06.2011 г.), замедления линейного роста (14.07.2011 г.), интенсивного роста в ширину и накопления сухой массы растения (28.08.2011 г.).
Определение эндогенных регуляторов роста проводили биологическим методом (по росту отрезков колеоптилей пшеницы сорта «Мироновская 808» в длину) с предварительной хроматографической разгонкой эфирных экстрактов [3]. При разгонке хроматограмм использовали щелочную смесь н-бутиловый спирт:аммиак:вода в соотношении 100:3:18. В качестве биотеста служили колеоптили пшеницы. Определяли свободную (физиологически активную) фракцию рострегулирующих веществ (ауксинов и ингибиторов) в материале (побеги дуба текущего года), фиксированном в парах кипящей смеси этанол:вода (1:1).
Результаты исследования и их обсуждение
Определение эндогенных регуляторов роста индольной (ауксины) и фенольной (ингибиторы роста) природы у клонов дуба черешчатого и колонновидного позволило выявить между ними существенные различия в содержании, динамике и балансе рострегулирующих веществ.
У обеих рамет (деревьев) в период интенсивных ростовых процессов в длину и по диаметру (май, июнь, август) выше уровень стимуляторов роста и их удельный вес (соотношение стимуляторы/ингибиторы) в общем балансе рострегулирующих веществ (табл. 1, рис. 1). У дуба черешчатого в этих побегах во все сроки исследования (май, июнь, июль, август) выше уровень и удельный вес ингибиторов роста.
Таблица 1
Соотношение стимуляторы/ингибиторы в общем балансе рострегулирующих веществ исходных деревьев (рамет) дуба колонновидного и черешчатого (в 2011 г.)
|
Дата взятия образца |
Соотношение стимуляторы/ингибиторы |
|
|
I-й прирост |
II-й прирост |
|
|
Дуб колонновидный, Кольцовский сквер |
||
|
24.05 |
1,56 ± 0,05 |
– |
|
16.06 |
1,66 ± 0,05 |
– |
|
14.07 |
0,79 ± 0,02 |
0,12 ± 0,004 |
|
28.08 |
1,96 ± 0,06 |
0,14 ± 0,004 |
|
Дуб колонновидный, улица Карла Маркса |
||
|
24.05 |
3,40 ± 0,10 |
– |
|
16.06 |
1,31 ± 0,04 |
– |
|
14.07 |
0,14 ± 0,004 |
0,42 ± 0,01 |
|
28.08 |
1,85 ± 0,06 |
0,83 ± 0,02 |
|
Дуб черешчатый, Семилуки |
||
|
24.05 |
0,08 ± 0,002 |
– |
|
16.06 |
0,08 ± 0,002 |
– |
|
14.07 |
0,30 ± 0,009 |
10,5 ± 0,32 |
|
28.08 |
0,32 ± 0,009 |
1,91 ± 0,06 |
Суммарное содержание ауксинов и ингибиторов роста (табл. 2) за исследованный отрезок онтогенеза (24.05–28.06) чётко отражает отмеченную выше закономерность: побеги дуба колонновидного первого прироста превалируют по содержанию (в 3,3 раза) и удельному весу (соотношение стимуляторы/ингибиторы в 7,9–9,8 раз) ауксинов; дуба черешчатого – по содержанию (в 2,4–5,4 раза) и удельному весу (в 2,3–2,8 раз) ингибиторов роста.
Дуб колонновидный, Кольцовский сквер
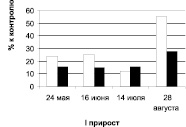
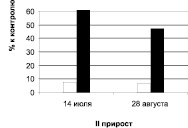
Дуб колонновидный, ул. Карла Маркса
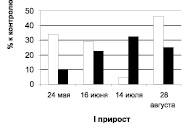
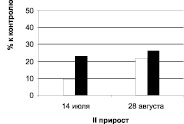
Дуб черешчатый, Семилукский питомник
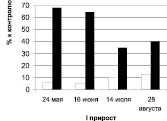
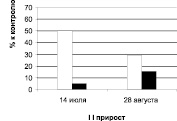
Рис. 1. Суммарное содержание рострегулирующих веществ в побегах исходных деревьев (рамет) дуба колонновидного и черешчатого, 2011 г.: белые столбики – стимуляторы, черные – ингибиторы
У побегов второго прироста наблюдается совершенно иная картина (табл. 1, 2, рис. 1): уровень стимуляторов и их удельный вес в общем балансе рострегулирующих веществ в побегах второго прироста выше у дуба черешчатого, а более высокий уровень и удельный вес ингибиторов отмечен у обеих рамет дуба колонновидного.
Клоны различаются по динамике содержания фитогормонов (рис. 2). Содержание ауксинов и их удельный вес в побегах первого прироста дуба колонновидного к моменту снижения интенсивности ростовых процессов в длину (июнь) падает и вновь резко возрастает к концу августа (период интенсивного роста по диаметру и накопления сухой массы). Направленность динамики содержания ауксинов обеих рамет практически идентична.
Динамика ингибиторов в побегах первого прироста дерева, растущего в Кольцовском сквере, аналогична в целом динамике их у дерева, растущего на улице Карла Маркса. Содержание ростингибирующих веществ резко возрастает к концу августа у первого дерева и уже к июлю – у второго, оставаясь практически на том же уровне в августе.
Динамика содержания регуляторов роста дуба черешчатого резко отличается от таковой дуба колонновидного. Содержание стимуляторов и их удельный вес в побегах первого прироста дуба черешчатого, находясь практически на одном уровне в мае и июне, возрастает к июлю и августу. Следует особо отметить низкий уровень и удельный вес ауксинов в побегах первого прироста, наблюдаемый на протяжении всего периода исследований (май – август), у этой формы дуба по сравнению с дубом колонновидным.
Уровень ингибиторов роста в мае и июне в побегах первого прироста дуба черешчатого представлен величинами, значительно превышающими уровень ауксинов.
Таблица 2
Суммарное содержание (средние данные) эндогенных рострегулирующих веществ в побегах дуба колонновидного и черешчатого за исследуемый отрезок онтогенеза (24.05–28.02.2011 г.) (% к контролю).
|
Место произрастания рамет |
Стимуляторы |
Ингибиторы |
Соотношение стимуляторы/ингибиторы |
|||
|
I-й прирост |
II-й прирост |
I-й прирост |
II-й прирост |
I-й прирост |
II-й прирост |
|
|
Дуб колонновидный |
||||||
|
Кольцовский сквер, Воронеж |
29,05 ± 0,87 |
7,00 ± 0,21 |
18,52 ± 0,56 |
54,20 ± 1,62 |
1,57 ± 0,05 |
0,13 ± 0,004 |
|
Улица Карла Маркса, Воронеж |
28,56 ± 0,86 |
15,68 ± 0,47 |
22,43 ± 0,67 |
24,63 ± 0,74 |
1,27 ± 0,04 |
0,64 ± 0,02 |
|
Дуб черешчатый |
||||||
|
Семилукский питомник, Воронежская область |
8,59 ± 0,26 |
39,80 ± 1,19 |
51,89 ± 1,56 |
10,07 ± 0,30 |
0,16 ± 0,005 |
3,96 ± 0,12 |
К июлю в отличие от дуба колонновидного содержание их резко снижается и снова возрастает в августе, оставаясь на протяжении всего периода исследований более высоким, чем уровень стимуляторов роста.
Определение фитогормонов в побегах второго прироста проведено в июле и августе (рис. 2). Выявлены изменения в их содержании во времени (от июля к августу) у обеих форм дуба. Как показывают данные, у побегов второго прироста разных рамет дуба колонновидного наблюдаются определённые различия в динамике рострегулирующих веществ. У дерева, растущего в Кольцовском сквере, количество стимуляторов роста остаётся в июле и августе на одном уровне и оно значительно ниже в оба срока, чем содержание ингибиторов, уровень которых к августу хотя и несколько снижается. У дерева, произрастающего на улице Карла Маркса, к августу повышается содержание и ингибиторов, и стимуляторов, но тем не менее уровень последних в оба срока также ниже, чем уровень ингибиторов роста.
У дуба черешчатого наблюдается противоположная картина: уровень стимуляторов и их удельный вес к августу падает, а ингибиторов возрастает. Причем, несмотря на то, что концентрация ауксинов снизилась к августу, их содержание в отличие от дуба колонновидного значительно превышает в оба периода уровень ингибиторов роста.
Полученные данные показывают, что физиолого-биохимические процессы, обусловливающие ростовую деятельность растения (в частности, образование и распределение ауксинов и ингибиторов) и выход его из состояния покоя, у дуба колонновидного начинаются несколько раньше, чем у дуба черешчатого.
Более высокий уровень и удельный вес рострегулирующих веществ, отмеченный выше в побегах первого прироста дуба колонновидного, может свидетельствовать о более высоком потенциале корнеобразования при культивировании их in vitro, в отличие от побегов второго прироста.
Высокий уровень и удельный вес ауксинов в общем балансе рострегулирующих веществ в побегах второго прироста дуба черешчатого свидетельствует о большей потенциальной способности к ризогенезу этих побегов.
Разгонка эфирных экстрактов рострегулирующих веществ, полученных из побегов клонов дуба колонновидного и черешчатого, на хроматограммах в смеси растворителей (н-бутиловый спирт:аммиак:вода) и просмотр их позволили отметить следующее.
Распределение зон стимулирования и ингибирования на хроматограммах у исследованных клонов различно. Для побегов первого прироста дуба колонновидного в целом характерен не только более высокий уровень, но и более широкий диапазон стимулирования; для дуба черешчатого – более широкий диапазон ингибирования. Во все сроки исследования у дуба черешчатого в этих побегах ингибиторы распределены практически почти по всему фронту хроматограмм.
а)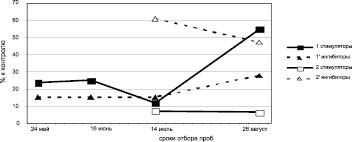
б)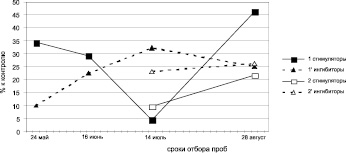
в)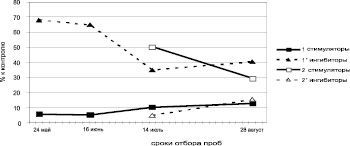
Рис. 2. Динамика содержания рострегулирующих веществ в побегах исходных деревьев (рамет) дуба колонновидного и черешчатого, 2011 г.: а – дуб колонновидный, Кольцовский сквер; б – дуб колонновидный, ул. Карла Маркса; в – дуб черешчатый, Семилукский питомник; 1 – I прирост, 2 – II прирост
У дуба колонновидного ростингибирующий эффект в период интенсивного роста в длину и по диаметру (май, июнь, август) проявляется обычно в двух-трёх зонах. И только в июле (период затухания, замедления роста побегов в длину) и здесь ингибиторы роста распределены практически по всему фронту хроматограмм.
В побегах второго прироста в оба срока исследований, наоборот, шире диапазон ингибирования у дуба колонновидного, а у дуба черешчатого – диапазон стимулирования. По всей видимости, клоны дуба черешчатого и колонновидного различаются между собой не только по количественному содержанию, но и по качественному составу рострегулирующих веществ.
Заключение
В результате проведенных исследований установлены различия в динамике и балансе эндогенных рострегулирующих веществ индольной (ауксины) и фенольной (ингибиторы роста) природы между клонами дуба черешчатого и колонновидного (пирамидального).
Физиолого-биохимические процессы (образование и распределение рострегулирующих веществ, их динамика), обусловливающие наряду с другими факторами ростовую деятельность растений в годичном цикле роста и его развития, у дуба колонновидного начинаются несколько раньше, чем у дуба черешчатого. Для дуба колонновидного характерен более высокий уровень и удельный вес ауксинов в побегах первого прироста, для дуба черешчатого – в побегах второго, что может в определённой степени свидетельствовать о разной потенциальной способности их к ризогенезу при культивировании in vitro. Кроме того, клоны дуба черешчатого и колонновидного различаются, по всей видимости, между собой не только по количественному содержанию, но и по качественному составу рострегулирующих веществ. Можно предположить, что при микроклональном размножении дуба колонновидного более целесообразно использовать побеги первого прироста, а дуба черешчатого – второго.
Рецензенты:
Свистова И.Д., д.б.н., профессор кафедры биологии растений и животных естественно-географического факультета ФГБОУ ВПО «Воронежский государственный педагогический университет», г. Воронеж;
Мелькумова Е.А., д.б.н., профессор кафедры ботаники, защиты растений, биохимии, микробиологии ФГБОУ ВПО «Воронежский государственный аграрный университет имени императора Петра I», г. Воронеж.
Работа поступила в редакцию 08.02.2013.
Библиографическая ссылка
Самсонова А.Е., Землянухина О.А., Карпеченко К.А., Калаев В.Н., Попов В.Н. ИЗУЧЕНИЕ РОСТРЕГУЛИРУЮЩИХ ВЕЩЕСТВ В ПОБЕГАХ, ИСХОДНЫХ ДЛЯ КУЛЬТИВИРОВАНИЯ in vitro, ДЕРЕВЬЕВ ДУБА КОЛОННОВИДНОГО И ЧЕРЕШЧАТОГО // Фундаментальные исследования. 2013. № 4-2. С. 388-393;URL: https://fundamental-research.ru/ru/article/view?id=31203 (дата обращения: 06.04.2026).