



Андродиэция (наличие у вида особей с тычиночными и особей с гермафродитными цветками) – довольно редкий вариант полового полиморфизма у цветковых растений [4]. В настоящее время мужская двудомность выявлена в разных семействах цветковых (Annonaceae, Malpighiaceae, Myrtaceae и др.). В умеренной зоне северного полушария андродиэция обнаружена в семействах Liliaceae, Ranunculaceae и Rosaceae [4, 11].
Geum urbanum L. – короткокорневищный кистекорневой гемикриптофит, поликарпик. Ареал вида западно-палеарктический, охватывает Европейскую часть России, Кавказ, Западную Сибирь, горы Средней Азии, Скандинавию, Среднюю и Атлантическую Европу, Гималаи, север Северной Африки [5]. Мезофит, встречается преимущественно в сосняках с примесью лиственных пород, реже в смешанных лесах и березняках, на опушках, в зарослях кустарников, довольно часто произрастает в садах и парках [6]. Крайне отрывочны сведения по экологии цветения и опыления G. urbanum [6, 14]. В связи с этим цель данной работы – выявление антэкологических особенностей G. urbanum.
Материалы и методы исследования
Материал собран в 2011–2014 г. в естественных условиях Московской области (Истринский район). Исследования проведены в фазу массового цветения вида (середина июня). Изучена морфология 50 обоеполых и тычиночных цветков G. urbanum. Цветение и опыление растений исследовали по методике А.Н. Пономарева [8] на пяти растениях. Для определения готовности рыльца воспринимать пыльцу применяли реакцию Робинсона [12]. Для определения видового состава собранных насекомых-опылителей и посетителей использован «Определитель…» [7].
Результаты исследования и их обсуждение
Цветки G. urbanum, как и у других представителей семейства Rosaceae [2, 3], не специализированы к определённым переносчикам пыльцы, что проявляется в их радиальной симметрии, отсутствию трубчатых частей (цветки открытого типа), раздельнолепестности венчика и раздельнолистности чашечки, а также собрании цветков в малоцветковые рыхлые синфлоресценции – дихазии.
Цветки гемициклические, актиноморфные, с двойным околоцветником, пятичленные. Гипантий (образован за счёт срастания оснований тычинок, лепестков и чашелистиков между собой) широкий и плоский. Чашечка из 5 свободных остающихся при плоде зелёных чашелистиков, с подчашием (образованным сросшимися попарно прилистниками соседних чашелистиков). Венчик свободнолепестный, из 5 жёлтых лепестков.
Андроцей многочленный, из 110–130 тычинок, расположенных в нескольких кругах на верхней, сильно опушённой стороне гипантия. Тычиночные нити длинные, тонкие. Пыльники качающиеся, овальной формы.
Гинецей апокарпный, из многочисленных спирально расположенных плодолистиков (200–220 шт.), собранных в округлую головку. Стилодии верхушечные, длинные, тонкие, в верхней трети с сочленением, при плодах с опадающим верхним сегментом и разрастающимся нижним. Нижняя часть стилодия с короткими щетинками, наверху крючковидно загнутая. Завязь яйцевидная, опушённая оттопыренными волосками.
Нектарники внутрицветковые. Нектароносная ткань у G. urbanum находится между андроцеем и гинецеем, выстилая внутреннюю часть гипантия. Кроме того, такая же ткань располагается между тычинками вокруг тычиночных нитей. Соседние тычинки оказываются настолько сближенными, что нектароносные участки ткани сливаются, образуя сплошной массив. Первый нектар появляется именно между тычинками различных кругов андроцея.
Размеры околоцветника и его частей у тычиночных цветков андромоно- и андродиэцичных особей G. urbanum несколько меньше, чем у обоеполых. Однако, структурных различий в строении чашечки, венчика и андроцея между обоеполыми и тычиночными цветками не наблюдается. Тычиночные цветки имеют редуцированный гинецей, представленный многочисленными зачаточными нефункционирующими плодолистиками, никогда не образующими плодиков. Нами не обнаружено окрашивания кончиков плодолистиков в тычиночных цветках раствором перманганата калия (реакция Робинсона) в течение всего периода цветения цветка. Степень редукции гинецея в тычиночных цветках достаточно сильно варьируется.
Пыльца. Зрелые пыльцевые зерна одиночные, 2-клеточные. Пыльцевые зерна 3-бороздно-оровые, эллипсоидальные, в очертании с полюса округлые, с экватора эллиптические. Длина полярной оси варьируется от 32,4 до 36,0 мкм, экваториальный диаметр – от 21,6 до 25,2 мкм. Фертильность пыльцы обоеполых и тычиночных цветков варьировалась от 54,7 до 100 %, составляя в среднем 92,8 и 94,3 % соответственно. Различий по степени фертильности пыльцы тычиночных и обоеполых цветков в нашем исследовании не выявлено.
Половая дифференциация. Впервые андродиэцию у G. urbanum обнаружил E. Loew [10]. Мужская двудомность у этого вида сопровождается андромоноэцией, при этом доля андромоноэцичных особей в популяциях может быть выше (до 10 %), чем особей только с тычиночными цветками [10]. В исследованной нами популяции наблюдалась такая же ситуация: доля андромоноэцичных особей в разные годы составляла в среднем 2–3 %, а доля растений с тычиночными цветками варьировалась от 0 до 0,8 %.
Суточная ритмика цветения. G. urbanum характеризуется утренней ритмикой раскрывания цветков (рисунок). Раскрывание цветков начинается около 5 ч утра, достигает максимума в 6–7 и заканчивается в 11–12 ч. Кривая суточного хода распускания носит одновершинный характер. Цветки раскрываются однократно (на ночь цветки не закрываются). По выявленным особенностям суточной ритмики цветения G. urbanum довольно близок к другим видам семейства Rosaceae [1].
Экология цветения. Продолжительность цветения обоеполых цветков G. urbanum равна двум дням (в жаркую погоду) или трём дням (в прохладную и пасмурную погоду). Обоеполые цветки G. urbanum слабо протогиничны. В плотных бутонах чашелистики соприкасаются своими краями и образуют свод над лепестками, тычинками с согнутыми тычиночными нитями и плодолистиками. В рыхлых бутонах, близких к раскрыванию, пыльники не вскрыты. В утренние часы в рыхлых бутонах рыльца внутренних плодолистиков начинают сильно окрашиваться раствором перманганата калия (созрели и готовы воспринимать пыльцу) и благородя быстрому росту стилодиев выдвигаются над лепестками. Когда цветок раскрывается, тычиночные нити изгибаются внутрь, так что пыльники располагаются близко к самым наружным плодолистикам. В это время все плодолистики в цветке созрели и восприимчивы к пыльце. При микроскопировании в ацетокармине на рыльцах видны немногочисленные прорастающие пыльцевые зерна. Рыльцевая стадия в функционировании обоеполого цветка продолжается весь первый день цветения и заканчивается только на второй день цветения.
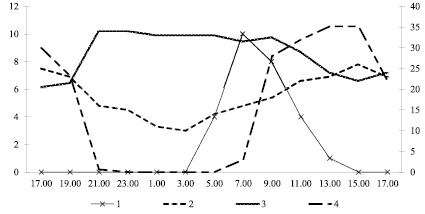
Суточная ритмика раскрывания цветков у Geum urbanum в Московской области. Условные обозначения: по оси Х – время наблюдений, ч; по левой оси Y – число раскрывшихся цветков, шт.; по правой оси Y – температура, °С (5 °С соответствует 10 % влажности и 12500 люкс). 1 – число раскрывшихся цветков, 2 – температура воздуха, 3 – влажность воздуха, 4 – освещённость
После полудня или ближе к вечеру первого дня самые дальние от центра цветка тычинки начинают изгибаться наружу, их пыльники вскрываются, и они поворачиваются вверх стороной, покрытой пыльцой. Таким образом, тычиночная стадия в развитии цветка также начинается в первый день. Из-за совмещения рыльцевой и тычиночной стадий возможна автогамия. Постепенно число пылящих тычинок увеличивается. В жаркую погоду к вечеру первого дня большинство рылец центральных плодолистиков уже утрачивает жизнеспособность.
На второй день цветения продолжается совмещение тычиночной и рыльцевой стадии в функционировании обоеполого цветка. С каждым часом в цветке все большее число тычиночных нитей выпрямляется, их пыльники вскрываются, тычинки самых наружных кругов заканчивают пыление. Пыльца самых внутренних тычинок почти всегда попадает на рыльца самых наружных плодолистиков, которые к этому времени ещё сохраняют способность воспринимать пыльцу.
Ксеногамия в обоеполых цветках G. urbanum обеспечивается в самом начале цветения цветков (в рыльцевую стадию развития) и посещения их насекомыми. При совмещении рыльцевой и тычиночной стадий (конец первого дня и второй день функционирования) наблюдается контактная автогамия в обоеполых цветках. Автогамия в цветках G. urbanum, по нашему мнению, играет достаточно большую роль, что связано с очень небольшим числом насекомых, которые посещают цветки данного вида и отвлекаются на более заметные цветки других цветущих растений. Гейтоногамия для G. urbanum не характерна в силу небольшого числа цветков в его синфлоресценции (дихазий) и разновременного их цветения в пределах особи. Таким образом, завязывание семян осуществляется преимущественно за счёт автогамии, которая обеспечивается наличием самосовместимости у G. urbanum [13]. По всей видимости, апомиктическое развитие семян у данного вида не встречается [14], в то время как апомиксис выявлен у близкородственного вида G. rivale [9].
У тычиночных цветков G. urbanum продолжительность жизни такая же, как у обоеполых. Рыльцевая фаза у них отсутствует. Развитие и функционирование андроцея происходит также, как описано у обоеполых цветков.
Экология опыления. На цветках G. urbanum нами зарегистрированы крайне немногочисленные насекомые трёх отрядов:
1. Coleoptera: Dasytes plumbeus Mull. (питаются нектаром, не опыляют цветков);
2. Diptera: Eristalis nemorum L., Melanostoma mellinum L. (питаются нектаром, не опыляют цветков);
3. Hymenoptera: Bombus terrestris L. (питаются нектаром и производят опыление цветков).
Крайне небольшой количественный и качественный состав насекомых-опылителей и посетителей цветков G. urbanum, по всей видимости, обусловлен значительно более низкой нектаропродуктивностью цветков в сравнении с одновременно цветущим близкородственным видом G. rivale. Тем не менее обоеполые цветки G. urbanum успешно завязывают семена. Так, в наших наблюдениях реальная семенная продуктивность варьировалась от 32 до 50 %.
Заключение
Отсутствие специализации к определённым насекомым у G. urbanum проявляется в целом комплексе признаков: цветки довольно крупные, открытого типа с легко доступным нектаром, радиально симметричные, с раздельнолепестным венчиком. Автогамия у G. urbanum обеспечивается непосредственным контактом функционирующих тычинок и плодолистиков из-за слабо выраженной протогинии, крайней немногочисленностью качественного и количественного состава насекомых-опылителей и посетителей цветков и самосовместимостью обоеполых цветков.
Рецензенты:
Викторов В.П., д.б.н., профессор, заведующий кафедрой ботаники МПГУ, г. Москва;
Шустов М.В., д.б.н., профессор, заведующий отделом флоры ГБС РАН, г. Москва.